Теги на стенах города: бескультурье или культура?
Подвид визуального уличного искусства — граффити — прочно укоренился в облике города. При этом качественных граффити в Севастополе по сей день не так уж много, а вот коротких подписей, или тегов, заполняющих фасады зданий даже на центральных улицах, превеликое множество, и они совсем не радуют глаз.
Все мы знаем о таких явлениях как граффити и стрит-арт, но также знаем и о вандализме. К чему отнести теги, которыми разрисованы не только стены дворов, но и фасады зданий на центральных улицах города?
Искусство «граффити» напрямую связано с субкультурой хип-хопа. Само по себе культурное направление «хип-хоп» появилось в Нью-Йорке в 80-е. Это движение развивалось в нескольких направлениях: музыкальное (рэп), танцевальное (брейк-данс) и изобразительное (граффити).
В этот период рисунки аэрозольной краской начинают появляться везде: на стенах дворов, в метро, вагонов поездов. Постепенно граффитчики начинают размечать «свою» территорию, оставлять свои подписи — теги. Появляются новые художественные стили, движения, художники начинают соревноваться: чем больше тегов-подписей, тем ты круче. Постепенно художники начинают рисовать «куски» — теги, нарисованные в замкнутом пространстве.
Появляются новые художественные стили, движения, художники начинают соревноваться: чем больше тегов-подписей, тем ты круче. Постепенно художники начинают рисовать «куски» — теги, нарисованные в замкнутом пространстве.
Такое самовыражение молодёжи можно было бы назвать безобидным, если бы «тегать» не начали повсюду. Сегодня в Севастополе можно столкнуться не только с мусором и свалками, но и с визуальным «шумом». Свои подписи молодые «художники» оставляют маркерами даже на фасадах зданий центральных улиц. Молодые люди «отметились» и на здании Драматического театра им. Лавренёва, и на фасадах зданий на Большой Морской улицы, в сквере на Историческом бульваре. И это только малая часть. Например, на проспекте Острякова орудуют «авто-вандалы».
Когда райтеры поймут, что разрисовывать гаражи и фасады исторических зданий — это разные вещи? В конце концов, должно быть хоть какое-то уважение к городу, в котором мы живём? Самое обидное, что идеально «замазать» неугодный тег невозможно. Ведь борются с такими художествами обычно белой краской, не подбирая оттенков. Потому на домах появляются пятна, а бывает и так, что тег «просвечивается» из-под краски.
Ведь борются с такими художествами обычно белой краской, не подбирая оттенков. Потому на домах появляются пятна, а бывает и так, что тег «просвечивается» из-под краски.
Тегают всё подряд: заборы, фасады домов, подъезды, гаражи, лестницы, вывески магазинов, фонари, телефонные будки, лавочки. Есть, конечно, и красивые, приятные взгляду граффити, но, увы, их немного. Хочется верить, что культура граффити будет подниматься на более высокую ступень и тёмные дворы и переулки увидят декоративные, трафаретные, рекламные, конкурсные граффити. Вписывающееся в ландшафт города граффити становится неотъемлемой его частью, знаковым местом в городе, на которое приходят посмотреть туристы и жители города.
Но пока того не случилось, хочется обратиться к райтерам с просьбой прекратить вандализм.
Работы известных в городе уличных художников можно посмотреть на нашем сайте: публиковались фоторепортажи с выставок Алексея Кислова и Павла Ртуе. Также можно ознакомиться с творчеством художницы Натальи Грезиной.
Кто такие теггеры и почему они уродуют город? (37 фото) — City4People_YKT
Многие горожане заметили как в последние годы хулиганы расписали различные городские поверхности своими граффити, а точнее «тегами». Тэги — это индивидуально разработанные подписи граффитчиков, их автографы и способ заявить о себе. Настенные росписи считаются самым распространенным видом граффити. Это движение зародилось задолго до появления аэрозольных баллончиков и маркеров.
На заре человечества, люди оставляли в пещерах отпечатки ладоней, имен, символов животных. Знакомая всем надпись “ДД.ММ.ГГГГ Здесь был Я” – тоже форма тэггинга, хотя сейчас такие тривиальные подписи встречаются всё реже.
Культура современного тэггинга зародилась в Америке 60-х годов, одновременно с появлением в продаже японских фломастеров. Одним из самых популярных тэггеров того времени стал некий Такi183. Работая курьером, он постоянно перемещался по городу и оставлял свою подпись на стенах, мусорных баках и лестницах.
Одним из самых популярных тэггеров того времени стал некий Такi183. Работая курьером, он постоянно перемещался по городу и оставлял свою подпись на стенах, мусорных баках и лестницах.
Прежде чем выходить на улицы, тэггер придумывает себе никнейм или же модифицирует школьную или детскую кличку. Это может быть не только псевдоним, многие создают комбинацию из любимых букв или цифр, которые несут в себе определенный смысл, например, номер дома или района, как у Такi183. Затем тэггер вырисовывает красивый шрифт, выходит на улицу и заявляет о себе – “я здесь, я есть, я в городе”.
Граффити сленг
All City King (Король всего города) — Райтер, рисующий по всему городу, получивший за это славуAll City (Ол сити) — Нанес тэги по всему городу, пометил весь город
Bomb/ Bombing — Бомба, бомбинг — нелегальное граффити
Buff (Бафф) — Удаление граффити со стен или поездов.Бафить (To buff) — закрашивать граффити
Sticker (Стикер) – Наклейка, на которой есть имя или рисунок человека, который его сделал. По сути это тот же тэг только он нанесён на наклейку
Stencil (Стенсил) — Трафарет для граффити и созданное с его помощью изображение
Tag (Тег) – Стилизованный автограф(подпись) художника (тэгнуть)
Taggers(Тэггеры) — Те, кто занимается только тэганием (нанесением никнеймов)
Writert (Райтер) – Граффити-художник
Каждый тэггер стремится к индивидуальности, чтобы его узнавали другие члены тусовки. Кто-то украшает надпись небольшими рисунками, кто-то предпочитает клеить тэги-стикеры, другие работают с помощью трафаретов. В каждой стране и городе, есть свои знаменитости, у которых большой стаж и своя философия. Абсолют для тэггера – получить среди своих звание «All city king», то есть «Король города».
Данное направление активно распространяется среди подростков.
А вы замечаете такие граффити в городе? Как думаете с этим можно бороться?
P. S. А вот так теггеры бравятся своими «подвигами» в твиттере. После нашего замечания ретвит был тут же удален
S. А вот так теггеры бравятся своими «подвигами» в твиттере. После нашего замечания ретвит был тут же удален
Присоединяйтесь к нашей команде Городских проектов в Якутске
Вместе мы сделаем наш город лучше!
Для этого надо всего лишь зарегистрироваться на сайте: russia.city4people.ru
И вступить в телеграм-чат отделения по ссылке в письме
ПОДПИСЫВАЙТЕСЬ НА НАС В СОЦСЕТЯХ:
Instagram @city4people_ykt
Twitter @city4people_ykt
Facebook @city4peopleYKT
Telegram-канал @urbanistika_severa
Граффити в Токио » BigPicture.ru
Пишет sanmai: После Осаки на улицах Токио бросается в глаза изобилие тегов и стикеров, одних из самых распространенных типов граффити.
Тег – это самый базовый вид граффити, состоящий обычно из одной лишь подписи райтера. Стикер-арт исполняет роль близкую по смыслу и применению к тегам, часто включают в себя и рукописную подпись. Рекомендую ознакомиться с терминологией современного граффити, на английском языке.
Стикер-арт исполняет роль близкую по смыслу и применению к тегам, часто включают в себя и рукописную подпись. Рекомендую ознакомиться с терминологией современного граффити, на английском языке.
Не могу сказать что тегов на улицах Осаки нет вообще – они есть, но вы можете пройти два километра в центре города и встретить только два неряшливых тега. В Токио вам сложно будет пройти сотню метров в центре города и не встретить ни одного тега или наклейки.
Смотрите также: Роботы в Токио, Современное искусство – граффити , Давка в токийском метро
(Всего 25 фото)
Спонсор поста: Отправить смс бесплатно в любую точку мира на телефон любого оператора.1. В японских городах много надземных переходов, к которым тут же прикреплены щиты с указаниями для автомобилистов. (Омотэсандо) 2. Столбы таких переходов также не остаются без внимания.3. На фоне – единственное в Японии кафе BVLGARI. 4. Редкий надземный переход в Токио не помечен хоть как-нибудь. 5. Видно что городские службы пытаются бороться с райтерами, но без особого успеха.

А вы знали, что у нас есть Instagram и Telegram?
Подписывайтесь, если вы ценитель красивых фото и интересных историй!
РАЗНИЦА МЕЖДУ ГРАФФИТИ И ТЕГАМИ | СРАВНИТЕ РАЗНИЦУ МЕЖДУ ПОХОЖИМИ ТЕРМИНАМИ — ЖИЗНЬ
Граффити и теги — это две формы уличного искусства, которые кажутся похожими, но между ними есть некоторая разница, и их не следует путать с одним и тем же. Стрит-арт сегодня получает признание в мире
Граффити против теговГраффити и теги — это две формы уличного искусства, которые кажутся похожими, но между ними есть некоторая разница, и их не следует путать с одним и тем же. Стрит-арт сегодня получает признание в мире как уникальный жанр. Граффити и теги — это две формы, которые очень популярны в городских городах большинства стран. Несмотря на то, что о них говорится в схожем контексте, это две разные формы. Под граффити понимаются надписи или рисунки на поверхности в общественном месте, тогда как тегами называют нанесение имени художника, подписи или логотипа на стене. Таким образом, тегирование считается очень простой формой граффити. В этой статье делается попытка выделить разницу между двумя формами, разъясняя, что подразумевается под каждой формой.
Таким образом, тегирование считается очень простой формой граффити. В этой статье делается попытка выделить разницу между двумя формами, разъясняя, что подразумевается под каждой формой.
Что такое граффити?
Граффити можно определить как любую картину, рисунок или написание чего-либо на поверхности в общественном месте. В большинстве стран это считается незаконным. Обычно граффити можно увидеть на стенах, мостах, метро и т. Д. Это обычно очень смелые и нетрадиционные презентации, которые идут рука об руку с хип-хоп культурой. Граффити бывает разных цветов, подчеркивая разнообразие стилей, а также различаются по размеру. Хотя некоторые виды граффити относительно небольшие, некоторые могут быть гигантскими по размеру и занимать большую площадь. Для их создания используются разные краски, например, аэрозольные баллончики. Граффити используется не только из-за своей эстетической привлекательности, но и для подчеркивания определенных социальных и политических проблем. Банды используют разные разновидности граффити для обозначения своей территории.
Что такое теги?
Маркировку можно определить как базовую форму граффити, когда писатель подписывает свое имя или подпись с помощью аэрозольной краски. В отличие от граффити, для создания художественного изображения которого требуется много навыков, тегирование не требует такого мастерства. Кроме того, для добавления тегов требуется ограниченное время. Так же, как и граффити, теги можно увидеть на стенах, улицах, остановках и т. Д. и может использоваться для обозначения территории. Это представление себя в общественном месте, а не абстрактное представление. Пометка также считается вандализмом по отношению к собственности, поскольку она портит поверхность. Это подчеркивает, что теги и граффити — это две разные формы.
Это подчеркивает, что теги и граффити — это две разные формы.
В чем разница между граффити и тегами?
• Граффити можно определить как любую картину, рисунок или письмо чего-либо на поверхности в общественном месте.
• Маркировка может быть определена как основная форма граффити, когда писатель подписывает свое имя или подпись с помощью аэрозольной краски.
• И граффити, и теги считаются незаконными.
• Добавление тегов — это основная форма граффити.
• В отличие от граффити, установка тегов требует ограниченного времени и навыков.
• В то время как тегирование — это больше представление о себе, граффити — это картина, имеющая очень много уровней, от самого искусства до выражения социально-политических проблем.
Изображения любезно предоставлены:
- Граффити Brocco (CC BY-SA 2.0)
- Пометка jaqian из Дублина, Ирландия (CC BY 2.0)
уличное искусство Владивостока выходит из подполья (ФОТО; ОПРОС) – Новости Владивостока на VL.
 ru
ruВ 2013 году еще несколько «монохромных» стен Владивостока «оделись в яркое», и процесс по облагораживанию серых городских ландшафтов с каждым годом только набирает обороты. О том, как андерграундное искусство выходит из подполья, за что платят граффитчикам и кто такие такие вандалы, чем «бомбы» отличаются от «тегов», а художники — от хулиганов, в интервью корреспонденту VL.ru рассказал руководитель проектов мастерской «33+1» Александр Бойко.
С наступлением тепла во Владивостоке вслед за деревьями и цветами, расцвели и стены: советские индустриальные мотивы появились на Инструментальном заводе, диггеры — на Партизанском проспекте, жизнерадостные «человечки» от художника Владимира Погребняка — в переходе на Молодежной, немыслимые клоуны — у городского цирка, моряки — на «1000 мелочей».
В мае городские власти и вовсе озадачили тех, кто считал граффити искусством протеста, царящем на урбанистических окраинах. Большая подпорная стена на Окатовой была целиком отдана для самовыражения местным художественным объединениям. При этом в росписи первой легальной площадки принимали участие команды «MIR», «ДВР», «HARM», и без того немало «наследившие» на всевозможных городских поверхностях.
При этом в росписи первой легальной площадки принимали участие команды «MIR», «ДВР», «HARM», и без того немало «наследившие» на всевозможных городских поверхностях.
Об уличном искусстве «по-владивостокски» корреспонденту VL.ru рассказал руководитель проектов Мастерской монументального искусства «33+1» Александр Бойко.
Судя по всему, во Владивостоке идет процесс выхода подпольного граффити-искусства из андерграунда на новый, легальный уровень, о чем говорит все увеличивающееся число «официально» расписанных стен. Так ли это?
Этот процесс был всегда, во всех странах. Просто у нас теперь стало возможно согласовывать арт-объекты. В России создается рынок уличного искусства, который на западе уже давно существует — когда люди могут этим профессионально заниматься, когда они востребованы. И после того, как рисовали граффити или делали какие-то нелегальные вещи, художники уже задумываются, заниматься мне этим профессионально и зарабатывать деньги, или пойти и стать инженером, юристом, врачом и зарабатывать там.
Кто эти люди, которые расписывают стены во Владивостоке? Откуда вы берете авторов?
Мы ищем людей, которые хотят как-то изменить внешний облик города. Если им это интересно, они рисуют сами на стенках или делают для них эскизы. Мы набираем каждый год сезонную группу художников — человек по 40. Их работа оплачивается. Многие остаются рисовать на 2-3 года.
У нас сейчас есть несколько постоянных талантливых авторов, которые с нами вместе выросли — это Кирилл Крючков, арт-группа «Мураед», Антон Эффект, Дмитрий Лучинин и Николай Кухаренко, Артем Здор, Алексей Новиков, и, конечно, группа «Конкрит Джангл».
Кто-то из них вышел из «подпольных» граффитчиков, уличных хулиганов?
Артем Здор и Алексей Новиков рисовали граффити. И они очень хорошие художники — могут нарисовать огромный фотореалистический портрет на всю стену. Правда, делают это в заброшенных зданиях, где никто не увидит. Кирилл Крючков — это вообще, наверное, один из самых первых граффити-художников Владивостока. Антон Эффект — тоже до сих пор рисует граффити.
Антон Эффект — тоже до сих пор рисует граффити.
Но они не вандалы. Если они рисуют граффити, это выглядит уместно. То есть, нельзя их работы встретить на только что отремонтированном здании. У них есть свои места, на которых только они рисуют и тренируются.
У уличных художников — своя идеология. Отказывался ли кто-то из андерграундных художников сотрудничать с вами из принципа не работать за деньги и по заказу?
Они все рады работать за деньги. Но многим никто не хочет платить, потому что зачастую они рисуют только буквы. Все граффитчики, которые более-менее умеют рисовать, — все занимаются оформлением интерьеров, татуировками и прочим-прочим. Чтобы кто-то отказывался работать — такого вообще ни разу не было. Все хотят работать и зарабатывать. Это мы отказываемся зачастую — не дотягивает уровень.
А что касается групп, которые часто вызывают негодование горожан — HARM, MIR, БУДНИ — вы как-то с ними работаете?
Нет, мы с ними не работаем. Много раз пытались их привлечь, но ничего не получается. Вот группа MIR — насколько я знаю, означает Made In Russia, — у них были неплохие шрифтовые композиции на Чуркине. Мы говорим не о так называемых «тегах» — когда баллоном или маркером оставляют какую-то подпись — это занимает много места и выглядит хуже всего. А если говорить о композиционных больших работах, то у MIR на Чуркине и на Спортивной были две хорошие работы, вписанные во всю стену, и это как-то облагораживало территорию, смотрелось уместно.
Много раз пытались их привлечь, но ничего не получается. Вот группа MIR — насколько я знаю, означает Made In Russia, — у них были неплохие шрифтовые композиции на Чуркине. Мы говорим не о так называемых «тегах» — когда баллоном или маркером оставляют какую-то подпись — это занимает много места и выглядит хуже всего. А если говорить о композиционных больших работах, то у MIR на Чуркине и на Спортивной были две хорошие работы, вписанные во всю стену, и это как-то облагораживало территорию, смотрелось уместно.
HARM делают классические «бомбы» — когда каким-то определенным образом пишут свое название. И чем недоступнее место, где они его оставили, тем круче.
Судя по всему эти ребята очень любят конспирацию, на контакт не идут…
Неизвестно, кто это, сколько их там людей. Никто не хочет сознаваться. Но все эти HARM, MIR и прочие-прочие, которые когда-то рисовали или сейчас рисуют — они сто процентов из одной тусовки, они все знакомы.
Но многие из них уже выросли, закончили институт, работают, некоторые имеют свой бизнес. И того задора, который был, когда они рисовали в 90-е, начале нулевых — его уже нет. Тогда на улице творился беспредел, а ты вроде бы культурный человек — ты рисуешь граффити, занимаешься уличным искусством, то есть ты не какой-то там наркоман, алкоголик. И на фоне общей разрухи в городе, в стране, это смотрелось круто.
Но на росписи стены на Окатовой, 18 эти группы тоже были?
Да, вроде были. Но Окатовая была не для того, чтобы кого-то поймать, и не для тех, кто делает «теги» и «бомбы». Эта стена отдана тем, кто хочет научиться рисовать на улице. Если ты делаешь «теги» — без проблем, ты можешь сделать их и на Окатовой, но толку от этого не будет, это никому неинтересно, это тебя не выделит никаким образом.
А если ты хочешь рисовать и тренироваться, делать росписи баллонами или кистями, и тебе нужно место, где это все увидят и при этом это будет легально и интересно, для этих людей создан такой объект как Окатовая, 18. И будут еще создаваться такие же. И это правильно, когда это как-то красиво оформлено, граффити — это хорошо.
И будут еще создаваться такие же. И это правильно, когда это как-то красиво оформлено, граффити — это хорошо.
Граффитчиков принято считать уличными вандалами. Можно и нужно ли с ними как-то бороться?
Граффитчики на сегодняшний день — это самые активные несогласные люди в обществе, потому что они смело выходят на улицу и пишут, что хотят, делают большие рисунки. Их просто надо любить и направлять в нужное русло. Это бесконечная проблема, которая не решена ни в одном городе. Не надо воевать с теми, кто рисует граффити, с ними надо дружить. Но «теги» — то, что загрязняет — будут появляться всегда. Всегда найдется молодой парень, который будет везде писать свое имя типа «Вася был здесь» — это тоже самое. Им важно, чтобы это видели, поэтому надписи будут на самых видных или необычных местах. Но они повторяют одно и то же — это все синонимы. А когда ты говоришь синонимами, ты не произносишь ничего.
Граффити — это только период подростковый, потому что он имеет большое количество ограничений. Если тебя это устраивает, то хорошо, если ты считаешь, что можешь что-то больше и лучше — тогда ты прошел этот этап.
Если тебя это устраивает, то хорошо, если ты считаешь, что можешь что-то больше и лучше — тогда ты прошел этот этап.
А среди ваших художников есть «хулиганы»?
Один раз мы поймали двух парней, которые рисовали во дворе граффити. Сначала они очень испугались, но мы им предложили сотрудничество. Сейчас они на первом курсе, но с нами лет с 15-16, заставляем их трудиться на благо общества. И это не мешает им везде оставлять «теги». Но на них есть возможность воздействовать — заставляем закрашивать. Сейчас они уже хорошо научились рисовать, интересно получается.
Однако, из тех, с кем мы работаем, я не знаю ни одного случая, чтобы они на видном месте что-то такое масштабное вандальное произвели, или нарисовали там, где это неуместно. Технология граффити — это нарисовал и ушел. Чтобы создать какую-то хорошую вещь, нужно много времени.
К вам не приходили из каких-нибудь «органов», организаций с просьбой сдать граффитчиков?
Было несколько раз. Приходили и по своим каналам просили найти, кто нарисовал что-то где-то. Но мы таким не занимаемся, нам это вообще неинтересно. Мы занимаемся искусством, а не сдачей мелких правонарушителей. Мы их не поощряем, но и не сдаем.
Приходили и по своим каналам просили найти, кто нарисовал что-то где-то. Но мы таким не занимаемся, нам это вообще неинтересно. Мы занимаемся искусством, а не сдачей мелких правонарушителей. Мы их не поощряем, но и не сдаем.
Ваши стены когда-нибудь подвергались вандализму?
На Феско-Холле, где нарисованы кумиры, часто рисуют маркером усы или пишут что-то националистическое. Но это очень мелко — какие-то школьники безобразничают, мы все исправляем.
А вот два года назад на подпорной стене на Молодежной с рельефами через месяц после того как мы закончили работу один человек нарисовал огромный член во всю длину рисунка, зайдя на все цвета. А там была ручная колеровка, вручную цвета подбирали. Хорошо, что у нас осталось немного красок. И мы исправили в тот же день. Но вот скажу, на Столетии есть пятиэтажки и со всех четырех сторон у них нарисован член во всю длину стен. И таких домов несколько. Видимо, старается один «художник», монументально и концептуально работает — очень узнаваемый стиль такой и больше ничего не рисует. И у граждан это вызывает недоумение.
И у граждан это вызывает недоумение.
А со стороны жителей города сталкивались с негативом по отношению к проектам и мастерам?
Конечно, не должно быть единого мнения. Любая роспись — это диалог, это вызов. Мы со своей стороны стараемся делать все максимально хорошо, но элементарно для какого-то человека это может быть не в его вкусе и он может подойти и сказать авторам, что ему не нравится. Обычно это неконструктивная критика и еще высказываются о том, что «украли много миллионов».
А еще со стороны комментаторов VL.ru — сталкивались. Если художник начинающий, он оформил какую-то стену, а тут такие негативные комментарии… Это может быть очень неприятно. Сначала я тоже думал: неужели мы действительно так плохо работаем? А потом я просто перестал читать комментарии на VL.ru, и все стало снова хорошо.
Опрос «УР»: граффити на стенах домов – вид творчества или акт вандализма?
«УР» узнал, что думают по поводу граффити и граффитчиков жители Екатеринбурга. В опросе приняли участие молодые люди от 18 до 34 лет.
В опросе приняли участие молодые люди от 18 до 34 лет.
– Граффити граффити рознь. Тупые теги, например, действительно портят внешний вид строений, но интересные, идейные, а главное, красивые граффити могут здорово украсить город.
Алексей Измайлов, 20 лет
– Отношусь негативно, если это связано с порчей общественного имущества. В Административном кодексе есть статья, которая позволяет привлечь к ответственности за подобные деяния. Но работает ли она, с этим не сталкивался.
Кирилл Савчук,34 года
– Мне сложно привести примеры, как у нас борются с вандализмом. Вроде есть уголовная статья, и штрафы предусмотрены, но ничего не меняется. Наверное, всем надо не закрывать глаза на действия вандалов, а останавливать их. Я так и делаю, когда вижу детей с баллончиками у заборов и жилых зон.
Диана Урмантаева, 24 года
– Наказывать нужно в зависимости от того, где граффитчики оставляют свои рисунки. Если это деревянный забор вокруг стройки, который потом снесут, то можно взыскать штраф в пользу строителей. Если это памятники культуры или другие достопримечательности, то надо давать за их порчу реальный срок от трех лет — чтобы у вандала было время обдумать свой варварский поступок.
Если это памятники культуры или другие достопримечательности, то надо давать за их порчу реальный срок от трех лет — чтобы у вандала было время обдумать свой варварский поступок.
Алексей Шалахов, 27 лет
– Если хулиганы изрисовали спинки сидений в общественном транспорте или подъездную дверь, то есть нанесли материальный вред, то это, конечно, плохо. Это вандализм, и такие поступки должны быть наказуемы. А вот граффити — совсем другое дело. Если граффитчиков не порицать, а привлекать к сотрудничеству для облагораживания общественных зон, где эти рисунки уместны, то город мог бы стать намного красочнее. Пример – стрит-арт на улице Толмачева в Екатеринбурге. А как бороться с тем, что творчеством точно не назовешь, затрудняюсь сказать. Но точно знаю, что каждому дорог его карман. То есть, нужно взимать штрафы в более крупном размере, чем сейчас.
Евгений Похольчик, 23 года
– Портить городское имущество – это ненормально. Вандализм должен наказываться по самому строгому счету. Не понимаю, почему кто-то считает, что может трогать то, что ему не принадлежит. Когда сталкиваюсь с тем, что кто-то из подростков исписывает мой подъезд, всегда делаю замечание. Если реакции на замечание не последует, то и полицией можно пригрозить.
Не понимаю, почему кто-то считает, что может трогать то, что ему не принадлежит. Когда сталкиваюсь с тем, что кто-то из подростков исписывает мой подъезд, всегда делаю замечание. Если реакции на замечание не последует, то и полицией можно пригрозить.
Марат Гиззатов, 18 лет
– Если граффити появляются на старых стенах, которые без них выглядели бы еще хуже, то я отношусь к этому терпимо. В целом же с настенной «живописью» надо бороться штрафами, не такое это ужасное преступление. При повторном нарушении можно увеличить штраф. Но если граффитчик упражнялся на памятниках или храмах, то условный срок или даже заключение до года было бы справедливым наказанием.
Алина Белоусова, 36 лет
– Один из методов борьбы с вандализмом – собрать жильцов своего дома и обсудить, как можно обезопасить его территорию, например, поставить забор. В новых жилых комплексах везде охраняемые дворы, а вот старые многоквартирные дома охранять некому, решать проблему придется самим жильцам. Ну, а за вандализм нужно наказывать штрафами, возможно и заключение в КПЗ.
Ну, а за вандализм нужно наказывать штрафами, возможно и заключение в КПЗ.
Опрос провела Алена Сорокина
Кто и зачем портит фасады зданий. Про каракули на стенах | AQUAdancing
Все мы видели несчастные черные каракули на стенах зданий. Изрисованы как старые здания, так и новые, недавно построенные здания обрастают паутиной из надписей.
Кто и зачем это делает?
Взрослым людям, с окрепшей и устоявшейся психикой гораздо комфортнее находится в гармоничной и чистой городской среде. Поэтому надписи такого плана раздражают. Если бы вы росли и жили где нибудь в гетто или на окраине провинциального города, то, разумеется, для вас это было бы нормой и вы бы даже не заметили, как на стене появляются тег за тегом.
Теги на новом зданииТеги на новом здании
Вы, скорее всего, понимаете, что оставляют эти каракули, называемые «тегами«, подростки и дети.
Подошёл к стене, нарисовал каракулю черным перманентным маркером — называется — тегнул.
Теги на электрическом щитеТеги на электрическом щите
Почему они тегают стены
Представляете, чем занимаются современные дети? Школа, секции, работа. Можно было бы организовать время ребенка так, чтоб не было времени на тегание стен, но не все взрослые могут занять своих детей делом не только из-за отсутствия времени, а иногда из-за материальных проблем.
Дети и подростки объединяются в компании и оставляют свой фирменный тег на стенах, где часто околачиваются, либо один подросток придумывает себе тег, исходя из клички или фамилии и рисует его где попало, типа: «Здесь был Вася», только тег должен быть максимально плохо читаем и понятен только своему автору.
Причины можно выделить следующие:
- Неблагоприятная среда обитания — когда живешь в грязи, то тег не считается чем то плохим.

- Отсутствие воспитания — родители с детства не объясняют, что такие рисунки это не только плохо, но и не красиво.
- Слишком много свободного времени — время растрачивается на гулянки с приченением ущерба городской среде.
- Соревновательный характер — каждый автор стремится оставить как можно больше тегов на большей площади, а может быть и затегать чужие подписи.
- Игра в ГТА Сан Андреас — там как раз была миссия, где надо было захватывать районы и оставлять свои теги.
Теги на гараже
Как бороться с тегами?
Полностью побороть теги не получится, локально можно устанавливать авторов по камерам, а потом заставлять смывать свои «произведения». Если установить автора одного тега, то можно найти ему очень много работы по удалению своего «творчества» со стен города.
Самый главный метод, это воспитание детей — ребенок сам не разберется, что красиво, а что нет. Нужно стараться создавать красоту вокруг детей и пояснять, что есть плохо и некрасиво. Нужно занимать время ребенка полезным и созидательным, чтоб он не занял его вредным и разрушительным.
Нужно стараться создавать красоту вокруг детей и пояснять, что есть плохо и некрасиво. Нужно занимать время ребенка полезным и созидательным, чтоб он не занял его вредным и разрушительным.
Всего доброго! Если статья понравилась — ставьте лайк или делайте репост.
Фрески с бирками Барри МакГи – Стены, покрытые сотнями красных бирок – Общественная доставка
Барри МакГи – Фрагмент росписи в Хьюстоне и Бауэри, Нью-Йорк, 2010 г. Twist) и Джош Ласкано (он же Amaze) нарисовали фреску на культовом углу Houston & Bowery в Нижнем Ист-Сайде Нью-Йорка, покрыв стену сотнями красных ярлыков, заполнив ее именами и командами разных авторов граффити. Впервые настенные росписи были заказаны в 2008 году, начиная с копии работы Кита Харинга 1982 года в ознаменование 50-летия покойного художника.В 2009 году стену расписала компания Os Gemeos, в 2010 году — Шепард Фейри, а вслед за Барри МакГи в 2010 году — Кенни Шарф, младший, Фейл, Ретна и другие. BY-NC 2. 0, автор carnagenycBarry McGee — Фреска в Хьюстоне и Бауэри, Нью-Йорк, 2010 г., фото: CC BY-NC 2.0, автор carnagenycBarry McGee — Фреска в Хьюстоне и Бауэри, Нью-Йорк, 2010 г., фото: CC BY-NC 2.0, автор Мэтью КраусБарри МакГи — Мурал на Хьюстоне и Бауэри, Нью-Йорк, 2010 г. Фото: CC BY-NC 2.0 Мэттью КраусБарри МакГи — Фреска Хьюстон и Бауэри, Нью-Йорк, 2010 г. Фото: CC BY-NC 2.0 Мэтью КраусБарри МакГи – Фреска Хьюстон и Бауэри, Нью-Йорк, 2010 г. Фото: CC BY-NC 2.0 Мэтью КраусБарри МакГи — Мурал на Хьюстоне и Бауэри, Нью-Йорк, 2010 г. Фото: CC BY-NC 2.0, автор Мэтью Краус. Мурал в Синдее был создан в 2011 году. Это сданных в эксплуатацию работ, состоящих из различных красных флажков.Чтобы иметь возможность прочитать некоторые теги, зрителям нужно было подойти вплотную к стене. Фреска достигала высоты более двух с половиной этажей. -2012, Tank Stream Way, фото: Джастин Топфер/Out of the Box ProjectsBarry McGee – Tag Mural в Сиднее (фрагмент), Австралия, 2011-2012, Tank Stream Way, фото: Жюстин Топфер/Out of the Box ProjectsБарри МакГи – Tag Мурал в Сиднее (фрагмент), Австралия, 2011–2012 гг.
0, автор carnagenycBarry McGee — Фреска в Хьюстоне и Бауэри, Нью-Йорк, 2010 г., фото: CC BY-NC 2.0, автор carnagenycBarry McGee — Фреска в Хьюстоне и Бауэри, Нью-Йорк, 2010 г., фото: CC BY-NC 2.0, автор Мэтью КраусБарри МакГи — Мурал на Хьюстоне и Бауэри, Нью-Йорк, 2010 г. Фото: CC BY-NC 2.0 Мэттью КраусБарри МакГи — Фреска Хьюстон и Бауэри, Нью-Йорк, 2010 г. Фото: CC BY-NC 2.0 Мэтью КраусБарри МакГи – Фреска Хьюстон и Бауэри, Нью-Йорк, 2010 г. Фото: CC BY-NC 2.0 Мэтью КраусБарри МакГи — Мурал на Хьюстоне и Бауэри, Нью-Йорк, 2010 г. Фото: CC BY-NC 2.0, автор Мэтью Краус. Мурал в Синдее был создан в 2011 году. Это сданных в эксплуатацию работ, состоящих из различных красных флажков.Чтобы иметь возможность прочитать некоторые теги, зрителям нужно было подойти вплотную к стене. Фреска достигала высоты более двух с половиной этажей. -2012, Tank Stream Way, фото: Джастин Топфер/Out of the Box ProjectsBarry McGee – Tag Mural в Сиднее (фрагмент), Австралия, 2011-2012, Tank Stream Way, фото: Жюстин Топфер/Out of the Box ProjectsБарри МакГи – Tag Мурал в Сиднее (фрагмент), Австралия, 2011–2012 гг. , Tank Stream Way, фото: Джастин Топфер/Out of the Box ProjectsBarry McGee — Tag Mural в Сиднее (частично полированный), Австралия, 2011–2012 гг., Tank Stream Way, фото: Жюстин Топфер/Out of the Box ProjectsСеребряный набросок Amaze и фреска с биркой Барри МакГи, Tank Stream Way, Сидней, Австралия, 2011–2012 гг. Уэй, Сидней, Австралия, 2011-2012 гг., фото: CC BY 2.0 от Emerald City to Oz
, Tank Stream Way, фото: Джастин Топфер/Out of the Box ProjectsBarry McGee — Tag Mural в Сиднее (частично полированный), Австралия, 2011–2012 гг., Tank Stream Way, фото: Жюстин Топфер/Out of the Box ProjectsСеребряный набросок Amaze и фреска с биркой Барри МакГи, Tank Stream Way, Сидней, Австралия, 2011–2012 гг. Уэй, Сидней, Австралия, 2011-2012 гг., фото: CC BY 2.0 от Emerald City to Oz
Местонахождение
Установлено в период с 23 сентября 2011 г. по 31 января 2012 г.
На улице Танк-Стрим, рядом с Питт-стрит, Сидней
Новый Южный Уэльс, 2000, Австралия
Теги Instagram собираются превратить его в вашу хромую стену Facebook
Эта статья из архива нашего партнера. Теперь, когда Instagram представил раздел «Фотографии с вами» для помеченных снимков, как и его материнская компания Facebook, вы можете забыть о красивых изображениях профессиональных фотографов и любителей, пытающихся быть профессионалами — ваше квадратное изображение мобильного телефона вот-вот превратится в отфильтрованное.
Facebook и Instagram уделяют большое внимание фотографиям. Исследование Гарвардской школы бизнеса показало, что 70% активности на Facebook связано с фотографиями. Марк Цукерберг сказал, представляя новую новостную ленту социальной сети, что Facebook будет делать акцент на больших изображениях, потому что половина существующих новостных лент занята постами с фотографиями. У него даже была диаграмма:
пользователей Facebook публикуют сотни миллионов фотографий каждый день — в канун Нового года и в первый день нового года Facebook достиг пика 1. 1 миллиард изображений, и вы можете поспорить, что на многих из них было столько же пьяных упивающихся людей, сколько и неодушевленных фейерверков. Instagram просматривает около 40 миллионов фотографий в день. Но изображения другого рода: некоторые из них красивы, в основном это тарелки для завтрака и очаровательные собаки.
1 миллиард изображений, и вы можете поспорить, что на многих из них было столько же пьяных упивающихся людей, сколько и неодушевленных фейерверков. Instagram просматривает около 40 миллионов фотографий в день. Но изображения другого рода: некоторые из них красивы, в основном это тарелки для завтрака и очаровательные собаки.
В этом особенность Instagram: он по-прежнему привлекает профессиональных фотографов и любителей, пытающихся сделать свои фотографии профессиональными. New York Times размещает фотографии из Instagram на своей первой странице. The New Yorker публикует их в своих блогах.Конечно, не все в Instagram является интеллектуальным, но большая часть из них, по крайней мере, пытается им быть, и даже селфи сами по себе являются своего рода жанром, более специфичным для платформы, чем реликвия с одноразовой камерой.
Facebook, с другой стороны, привлекает фотографии, которые могут привести к увольнению с работы. Поиск по фразе «Фото уволили с работы в Facebook» выдает не менее 1,5 миллиона таких кошмаров. Facebook — это свалка фотографий безрассудной развязности, рай для фотографий мобильных телефонов без фильтров — в прямом или переносном смысле.Там нет художественной панели, как в Instagram, где вы, по крайней мере, пытаетесь произвести впечатление на людей своей фотографией, а не просто пытаетесь показать, с кем вы проводите время. Существующие теги @ в Instagram — это почти счастливая боковая панель: сколько имен ваших друзей в Instagram вы знаете наизусть и насколько они вам небезразличны?
Facebook — это свалка фотографий безрассудной развязности, рай для фотографий мобильных телефонов без фильтров — в прямом или переносном смысле.Там нет художественной панели, как в Instagram, где вы, по крайней мере, пытаетесь произвести впечатление на людей своей фотографией, а не просто пытаетесь показать, с кем вы проводите время. Существующие теги @ в Instagram — это почти счастливая боковая панель: сколько имен ваших друзей в Instagram вы знаете наизусть и насколько они вам небезразличны?
Вот где вступает в игру психология тегов Facebook: «Люди любят отмечать своих друзей и семью», — говорится в пресс-релизе Facebook от 2010 года.И это правда: люди любят тратить время на просмотр фотографий в Facebook из-за друзей, заклятых врагов, бывших или влюбленных, отмеченных на них. Вместо того, чтобы покорить нас фильтрами и субботними вечерними закатами, Facebook покорил нас людьми. Этим и интересны картинки. Это также сделало просмотр картинок относительно приемлемой формой молчаливого социального взаимодействия. Вкладка «Фотографии» в профиле Facebook в наши дни является известным центром, где можно узнать все о человеке. Теги дали Facebook и его фотографиям определенную цель для существования, для знакомства с кем-то — в лучшую или в худшую сторону.
Вкладка «Фотографии» в профиле Facebook в наши дни является известным центром, где можно узнать все о человеке. Теги дали Facebook и его фотографиям определенную цель для существования, для знакомства с кем-то — в лучшую или в худшую сторону.
Но Instagram превратился во что-то без всего этого. Это никогда не было о кто. Прямо сейчас мой стрим в Instagram — это фотографии разных вещей — пейзажей, достопримечательностей, моды — всего несколько кадров с настоящими человеческими лицами; на момент написания этой статьи на вкладке «Популярное» в Instagram были только фотографии Эллен ДеДженерес и Милы Кунис. Конечно, с тегами в Instagram все изменится. Люди начнут думать о других людях. И скоро Instagram станет Facebook с фильтрами.Спорим, ты не можешь ждать, верно?
Эта статья из архива нашего партнера The Wire .
Границы | Флуоресцентные метки для изучения структуры и динамики клеточных стенок
Введение
В настоящее время мы достаточно хорошо понимаем структуру полисахаридов, и, во многом благодаря использованию генетики, происходит быстрый прогресс в понимании механизмов и регуляции синтеза стенок. Основной задачей на будущее будет понимание архитектуры клеточной стенки и ее динамики во время роста.Здесь мы рассмотрим недавний прогресс в области флуоресцентной микроскопии, который должен в значительной степени способствовать достижению этой цели.
Основной задачей на будущее будет понимание архитектуры клеточной стенки и ее динамики во время роста.Здесь мы рассмотрим недавний прогресс в области флуоресцентной микроскопии, который должен в значительной степени способствовать достижению этой цели.
Структура и динамика клеточной стенки
Клеточные стенки растений представляют собой высокодинамичные и гетерогенные структуры, которые различаются между типами клеток, стадиями роста, а также между микродоменами в одной клеточной стенке. Необходимы инструменты для визуализации этой неоднородности в живых клетках с высоким пространственным и временным разрешением.
Методы электронной микроскопии [просвечивающая электронная микроскопия (ПЭМ) и сканирующая электронная микроскопия с полевой эмиссией (FESEM)] сыграли важную роль в получении информации об архитектуре стен.Однако для этих методов требуется фиксированный материал, который не всегда репрезентативен для нативных гидратированных клеточных стенок и дает мало информации о динамике клеточных стенок или вообще не дает ее. Использование флуоресцентных зондов должно обеспечить доступ к динамическим аспектам. Доступно большое количество антигликановых антител (Knox, 2008; Moller et al., 2008; Pattathil et al., 2010; Ralet et al., 2010, см. обзор Lee et al., 2011; PlantProbes CarboSource Services). . Кроме того, растущее число секвенированных геномов растений и микроорганизмов обеспечивает практически неограниченный запас зачастую высокоспецифичных доменов, связывающих углеводы (Guillen et al., 2010). Эти зонды, как правило, дают статическую картину, но также возможна маркировка живых клеток. Например, Домозич и соавт. (2009) использовали антитела для изучения созревания пектина у одноклеточной харофитовой водоросли Penium margaritaceum . Будет интересно изучить аналогичные подходы к изучению динамики стенок у высших растений. При использовании этих зондов требуется осторожность, поскольку полисахаридные эпитопы часто маскируются другими полимерами (Marcus et al., 2008, 2010).
Использование флуоресцентных зондов должно обеспечить доступ к динамическим аспектам. Доступно большое количество антигликановых антител (Knox, 2008; Moller et al., 2008; Pattathil et al., 2010; Ralet et al., 2010, см. обзор Lee et al., 2011; PlantProbes CarboSource Services). . Кроме того, растущее число секвенированных геномов растений и микроорганизмов обеспечивает практически неограниченный запас зачастую высокоспецифичных доменов, связывающих углеводы (Guillen et al., 2010). Эти зонды, как правило, дают статическую картину, но также возможна маркировка живых клеток. Например, Домозич и соавт. (2009) использовали антитела для изучения созревания пектина у одноклеточной харофитовой водоросли Penium margaritaceum . Будет интересно изучить аналогичные подходы к изучению динамики стенок у высших растений. При использовании этих зондов требуется осторожность, поскольку полисахаридные эпитопы часто маскируются другими полимерами (Marcus et al., 2008, 2010).
Недавно также был достигнут значительный прогресс в визуализации динамики целлюлозы в живых клетках с использованием специфического для целлюлозы красителя Pontamine Fast Scarlet 4B (S4B; Anderson et al. , 2010). Конфокальная микроскопия окрашенных S4B корней выявила ориентацию микрофибрилл целлюлозы. Интересно, что окрашивание не мешало росту, и во время растяжения клеток можно было наблюдать вращение микрофибрилл от поперечного к продольному к оси растяжения. Это первая in vivo проверка гипотезы «многосетевого роста» 57-летней давности (Roelofsen and Houwink, 1953).
, 2010). Конфокальная микроскопия окрашенных S4B корней выявила ориентацию микрофибрилл целлюлозы. Интересно, что окрашивание не мешало росту, и во время растяжения клеток можно было наблюдать вращение микрофибрилл от поперечного к продольному к оси растяжения. Это первая in vivo проверка гипотезы «многосетевого роста» 57-летней давности (Roelofsen and Houwink, 1953).
Ферментативная активность также может быть визуализирована в стенке с использованием флуоресцентных донорских субстратов.Сульфородаминовые конъюгаты олигосахаридов ксилоглюкана (XGO-SR) использовали для визуализации активности эндогенной ксилоглюкан-эндо-трансгликозилазы (XET) (энзима ксилоглюкан-трансгликозилазы/гидролазы, XTH) в арабидопсисе и корнях табака и культивируемых клетках (Vissenberg et al., 2005), а также в древесине тополя (Nishikubo et al., 2011). XGO-SR проникают в ткани и включаются в ксилоглюкан в клеточной стенке, вызывая оранжевую флуоресценцию, указывающую на одновременную колокализацию в одном и том же компартменте активных цепей XET и акцепторных ксилоглюкановых цепей. Интересно, что фибриллярный паттерн, связанный с микрофибриллами целлюлозы, наблюдался в удлиняющихся клетках, указывая на то, что XTH действуют на ксилоглюканы, прикрепленные к микрофибриллам целлюлозы. Активность XET, обнаруживаемая в удлиняющихся клетках, может играть роль во включении вновь синтезированных молекул ксилоглюкана в клеточную стенку и/или ремоделировании существующей сети целлюлоза/ксилоглюкан. Активность гликозилгидролазы также можно визуализировать in situ . Например, активность эндо-β-1,4-ксилоглюканазы была обнаружена в muro с использованием резоруфина β-гликозида ксилоглюкоолигосахарида (XXXG-β-Res; Ibatullin et al., 2009) и активность целлюлазы с использованием резоруфинил-β-D-целлобиозида (Glc-2-β-Res; Takahashi et al., 2009).
Интересно, что фибриллярный паттерн, связанный с микрофибриллами целлюлозы, наблюдался в удлиняющихся клетках, указывая на то, что XTH действуют на ксилоглюканы, прикрепленные к микрофибриллам целлюлозы. Активность XET, обнаруживаемая в удлиняющихся клетках, может играть роль во включении вновь синтезированных молекул ксилоглюкана в клеточную стенку и/или ремоделировании существующей сети целлюлоза/ксилоглюкан. Активность гликозилгидролазы также можно визуализировать in situ . Например, активность эндо-β-1,4-ксилоглюканазы была обнаружена в muro с использованием резоруфина β-гликозида ксилоглюкоолигосахарида (XXXG-β-Res; Ibatullin et al., 2009) и активность целлюлазы с использованием резоруфинил-β-D-целлобиозида (Glc-2-β-Res; Takahashi et al., 2009).
Другим крупным достижением стало использование клик-химии для изучения метаболизма гликанов in vivo . Стратегия основана на метаболическом включении азидных сахаров в молекулы-мишени, подготавливая их к селективной ковалентной реакции с флуоресцентными зондами. Азид имеет небольшие размеры, стабилен в биологических системах и избирательно реагирует с фосфинами или активированными алкинами.Ранее лигирование Штаудингера использовалось для мечения гликоконъюгатов, меченных азидо-сахаром, на культивируемых клетках и у живых мышей (Prescher et al., 2004). Катализируемая медью клик-химия показала включение азидосодержащего аналога фукозы в гликопротеины посредством пути утилизации фукозы в клетках jurkat (Sawa et al., 2006). После этих первых экспериментов метод получил дальнейшее развитие и был адаптирован для клеток растений. Андерсон и др. (2012) использовали алкин фукозы (FucAI) для мечения пектинового рамногалактуронана-I (RG-I) в корнях проростков Arabidopsis .Используя эксперименты с импульсной погоней, они смогли визуализировать отложение пектина и переориентацию пектиновой сети во время удлинения клеток эпидермального корня. Проблема с этим методом заключается в том, что составы катализаторов Cu(I) токсичны, что препятствует их использованию в живых клетках.
Азид имеет небольшие размеры, стабилен в биологических системах и избирательно реагирует с фосфинами или активированными алкинами.Ранее лигирование Штаудингера использовалось для мечения гликоконъюгатов, меченных азидо-сахаром, на культивируемых клетках и у живых мышей (Prescher et al., 2004). Катализируемая медью клик-химия показала включение азидосодержащего аналога фукозы в гликопротеины посредством пути утилизации фукозы в клетках jurkat (Sawa et al., 2006). После этих первых экспериментов метод получил дальнейшее развитие и был адаптирован для клеток растений. Андерсон и др. (2012) использовали алкин фукозы (FucAI) для мечения пектинового рамногалактуронана-I (RG-I) в корнях проростков Arabidopsis .Используя эксперименты с импульсной погоней, они смогли визуализировать отложение пектина и переориентацию пектиновой сети во время удлинения клеток эпидермального корня. Проблема с этим методом заключается в том, что составы катализаторов Cu(I) токсичны, что препятствует их использованию в живых клетках. Для преодоления этой проблемы были разработаны различные подходы. Сориано Дель Амо и др. (2010) сообщили, что BTTES, лиганд Cu(I) на основе трис(триазолилметил)амина, быстро способствует реакции циклоприсоединения в живых системах без явной токсичности.Этот катализатор впервые позволил неинвазивно визуализировать фукозилированные гликаны во время раннего эмбриогенеза рыбок данио. Недавние альтернативные подходы основаны на «клик-химии без Cu» (обзор см. в Chang et al., 2010; Jewett and Bertozzi, 2010) для увеличения скорости циклоприсоединения без необходимости использования катализатора. Эти методы использовались для мечения биомолекул у рыбок данио (Laughlin et al., 2008) и у живых мышей (Chang et al., 2010). Крайне интересно будет использовать подобные методы для визуализации мест синтеза, депонирования и оборота различных полисахаридов в растениях (Wallace, Anderson, 2012).
Для преодоления этой проблемы были разработаны различные подходы. Сориано Дель Амо и др. (2010) сообщили, что BTTES, лиганд Cu(I) на основе трис(триазолилметил)амина, быстро способствует реакции циклоприсоединения в живых системах без явной токсичности.Этот катализатор впервые позволил неинвазивно визуализировать фукозилированные гликаны во время раннего эмбриогенеза рыбок данио. Недавние альтернативные подходы основаны на «клик-химии без Cu» (обзор см. в Chang et al., 2010; Jewett and Bertozzi, 2010) для увеличения скорости циклоприсоединения без необходимости использования катализатора. Эти методы использовались для мечения биомолекул у рыбок данио (Laughlin et al., 2008) и у живых мышей (Chang et al., 2010). Крайне интересно будет использовать подобные методы для визуализации мест синтеза, депонирования и оборота различных полисахаридов в растениях (Wallace, Anderson, 2012).
Флуоресцентная микроскопия для изучения в живых клетках внутриклеточной динамики и стехиометрии белковых комплексов, участвующих в биосинтезе клеточной стенки
Технологии лазерной сканирующей конфокальной микроскопии (LSCM) и конфокальной микроскопии с вращающимся диском (SDCM) использовались для изучения, часто очень подробно, динамики белков, участвующих в синтезе клеточной стенки. В то время как LSCM использует одну точечную диафрагму для оптического сечения, SDCM использует массив точечных отверстий возбуждения и излучения (можно использовать одну или две матрицы точечных отверстий) на быстро вращающемся диске, так что массив точечных отверстий осматривает все поле зрения более 1000 раз. /с.Высокая скорость сканирования не только повышает скорость получения изображений (до 360 кадров/с), но и снижает пиковую плотность возбуждающего света до нескольких мкВт/мкм 2 , тем самым повышая эффективность флуоресценции и уменьшая фотообесцвечивание и эффекты фотоповреждения по сравнению с точечным сканированием. Возможно, самое главное, поскольку все конфокальное поле зрения может быть захвачено малошумящей камерой с высокой квантовой эффективностью вместо фотоумножителя (ФЭУ), системы SDCM имеют более чем 50-кратное увеличение эффективности захвата света (т. .е., среднее количество фотонов, полученных от одной гранулы, нанесенное на график в зависимости от индекса экспозиции, который определяется путем измерения скорости фотообесцвечивания, а не фактической флуоресценции в каждом анализируемом месте образца), что приводит к увеличению отношения сигнал/шум в несколько раз.
В то время как LSCM использует одну точечную диафрагму для оптического сечения, SDCM использует массив точечных отверстий возбуждения и излучения (можно использовать одну или две матрицы точечных отверстий) на быстро вращающемся диске, так что массив точечных отверстий осматривает все поле зрения более 1000 раз. /с.Высокая скорость сканирования не только повышает скорость получения изображений (до 360 кадров/с), но и снижает пиковую плотность возбуждающего света до нескольких мкВт/мкм 2 , тем самым повышая эффективность флуоресценции и уменьшая фотообесцвечивание и эффекты фотоповреждения по сравнению с точечным сканированием. Возможно, самое главное, поскольку все конфокальное поле зрения может быть захвачено малошумящей камерой с высокой квантовой эффективностью вместо фотоумножителя (ФЭУ), системы SDCM имеют более чем 50-кратное увеличение эффективности захвата света (т. .е., среднее количество фотонов, полученных от одной гранулы, нанесенное на график в зависимости от индекса экспозиции, который определяется путем измерения скорости фотообесцвечивания, а не фактической флуоресценции в каждом анализируемом месте образца), что приводит к увеличению отношения сигнал/шум в несколько раз.
Используя SDCM, можно было наблюдать флуоресцентные точки в плазматической мембране клеток гипокотиля Arabidopsis , экспрессирующих XFP, слитый с различными компонентами целлюлозосинтазного комплекса (CSC; CESA3, CESA6, CESA5 или CSI1/POM2; Paredez et al., 2006; Деспре и др., 2007; Гу и др., 2010; Бишофф и др. , 2011; Брингманн и др., 2012 г.; Санчес-Родригес и др., 2012). Эти точки мигрируют в обоих направлениях по линейным траекториям и, скорее всего, соответствуют отдельным CSC. Поскольку CSCs продвигаются полимеризацией цепей глюкана, скорость CSCs может быть использована для количественной оценки синтеза целлюлозы in vivo . Эти инструменты широко использовались для изучения роли кортикальных микротрубочек в управлении траекториями РСК, их встраивании в плазматическую мембрану, регуляции движения РСК путем фосфорилирования, а также действия малых ингибиторов биосинтеза целлюлозы (DeBolt et al., 2007а,б; Кроуэлл и др., 2009 г.; Гутьеррес и др., 2009 г.; Чен и др., 2010 г.; Бишофф и др., 2011).
, 2011; Брингманн и др., 2012 г.; Санчес-Родригес и др., 2012). Эти точки мигрируют в обоих направлениях по линейным траекториям и, скорее всего, соответствуют отдельным CSC. Поскольку CSCs продвигаются полимеризацией цепей глюкана, скорость CSCs может быть использована для количественной оценки синтеза целлюлозы in vivo . Эти инструменты широко использовались для изучения роли кортикальных микротрубочек в управлении траекториями РСК, их встраивании в плазматическую мембрану, регуляции движения РСК путем фосфорилирования, а также действия малых ингибиторов биосинтеза целлюлозы (DeBolt et al., 2007а,б; Кроуэлл и др., 2009 г.; Гутьеррес и др., 2009 г.; Чен и др., 2010 г.; Бишофф и др., 2011).
Внутриклеточный перенос и время жизни белков удобно изучать с помощью фотоактивируемых или фотоконвертируемых флуоресцентных белков (Patterson et al., 2010). Фотоактивируемые флуоресцентные белки использовались в растениеводстве для анализа динамической взаимосвязи между ER и стопками Гольджи в эпидермальных клетках листьев табака (Runions et al. , 2006) или распределения каналов KAT1 K + в листьях табака (Sutter et al. др., 2006). Свойство фотоконвертируемых флуоресцентных белков демонстрировать ярко выраженные спектральные изменения, индуцированные светом, позволяет анализировать перенос белков с помощью импульсного анализа. Например, УФ-индуцированная фотоконверсия mEosFP из зеленого в красный цвет использовалась для изучения перемещения PIN2-mEosFP из одного внутриклеточного местоположения в другое (Dhonukshe et al., 2007). Такие инструменты будут очень полезны для анализа перемещения компонентов CSC или других ферментов биосинтеза клеточной стенки.
, 2006) или распределения каналов KAT1 K + в листьях табака (Sutter et al. др., 2006). Свойство фотоконвертируемых флуоресцентных белков демонстрировать ярко выраженные спектральные изменения, индуцированные светом, позволяет анализировать перенос белков с помощью импульсного анализа. Например, УФ-индуцированная фотоконверсия mEosFP из зеленого в красный цвет использовалась для изучения перемещения PIN2-mEosFP из одного внутриклеточного местоположения в другое (Dhonukshe et al., 2007). Такие инструменты будут очень полезны для анализа перемещения компонентов CSC или других ферментов биосинтеза клеточной стенки.
Восстановление флуоресценции после фотообесцвечивания (FRAP) и потеря флуоресценции при фотообесцвечивании (FLIP) являются дополнительными инструментами для визуализации белков клеточной стенки.Используя FRAP, частота доставки комплексов CESA к плазматической мембране была оценена примерно в 5 событий/мкм 2 ч (Gutierrez et al., 2009). В этом случае короткий период обесцвечивания, за которым следуют измерения после обесцвечивания, используется для измерения любого восстановления флуоресценции в обесцвеченной области неотбеленными частицами CESA. FLIP был использован для изучения распределения CSCs в развивающихся сосудах ксилемы. Под конфокальным микроскопом желтый флуоресцентный белок (YFP)-CSC выглядит как поперечные полосы в развивающихся сосудах ксилемы (Wightman and Turner, 2008).Чтобы преодолеть проблему высокой плотности YFP-CSC и глубины фокуса, необходимой для визуализации более глубоких слоев органа, использовали FLIP. При таком подходе измерялось движение обесцвеченных частиц наружу от обесцвеченной области (White and Stelzer, 1999). Одинаковая скорость потери флуоресценции по обе стороны от обесцвеченной зоны свидетельствует о том, что эти CSC, как и CSC первичной клеточной стенки, также мигрируют в обоих направлениях. Подвижность комплексов YFP-CSC была оценена примерно в 7 мкм/с, что намного выше, чем ранее сообщалось для CSC первичной стенки.Однако маловероятно, чтобы такая высокая скорость отражала скорость синтеза целлюлозы (которая составляла бы около 14 000 остатков глюкозы/цепь против 10 остатков/цепь для первичных клеточных стенок).
FLIP был использован для изучения распределения CSCs в развивающихся сосудах ксилемы. Под конфокальным микроскопом желтый флуоресцентный белок (YFP)-CSC выглядит как поперечные полосы в развивающихся сосудах ксилемы (Wightman and Turner, 2008).Чтобы преодолеть проблему высокой плотности YFP-CSC и глубины фокуса, необходимой для визуализации более глубоких слоев органа, использовали FLIP. При таком подходе измерялось движение обесцвеченных частиц наружу от обесцвеченной области (White and Stelzer, 1999). Одинаковая скорость потери флуоресценции по обе стороны от обесцвеченной зоны свидетельствует о том, что эти CSC, как и CSC первичной клеточной стенки, также мигрируют в обоих направлениях. Подвижность комплексов YFP-CSC была оценена примерно в 7 мкм/с, что намного выше, чем ранее сообщалось для CSC первичной стенки.Однако маловероятно, чтобы такая высокая скорость отражала скорость синтеза целлюлозы (которая составляла бы около 14 000 остатков глюкозы/цепь против 10 остатков/цепь для первичных клеточных стенок). Более вероятно, что авторы измерили гораздо более быстрое движение внутриклеточных компартментов, содержащих CSC (Wightman et al., 2009).
Более вероятно, что авторы измерили гораздо более быстрое движение внутриклеточных компартментов, содержащих CSC (Wightman et al., 2009).
Флуоресцентная микроскопия также широко используется для изучения белок-белковых взаимодействий в растениях. Используемые методы включают перенос энергии флуоресцентного резонанса (FRET) или бимолекулярную флуоресценцию (BiFC) или комплементацию люциферазы (Jares-Erijman and Jovin, 2003; Bracha-Drori et al., 2004; Фудзикава и Като, 2007 г.). Стратегии BiFC использовались для определения специфических взаимодействий между различными изоформами CESA (Desprez et al., 2007; Timmers et al., 2009). N-концевая половина YFP слита с первой изоформой CESA, а С-концевая — со второй изоформой. Обнаружение сигнала YFP указывает на события белок-белкового взаимодействия, поскольку половинки YFP находятся в достаточной близости, чтобы сформировать интактную молекулу YFP. Это простой и быстрый метод, однако важно отметить, что он создает необратимые взаимодействия, и необходимо включить соответствующие элементы управления, такие как белки, присутствующие в одном компартменте, которые не взаимодействуют с тестируемым белком.
Недавнее усовершенствование в обнаружении молекул на поверхности растительных клеток связано с использованием флуоресцентной микроскопии полного внутреннего отражения (TIRFM). Этот метод основан на физическом явлении полного внутреннего отражения (ПВО), которое возникает, когда луч света падает на границу между двумя материалами с разными показателями преломления ( n ), а угол падения больше критического угла падения. . В этих условиях весь свет отражается обратно в среду с более высоким значением n .При ПВО на границе формируется световая волна ближнего поля; эта «затухающая волна» (ЭВ) может проникать в поверхность среды на глубину, примерно равную 1/3 длины волны падающего света. TIRFM может обеспечить около 100 нм объема возбуждения вблизи плазматической мембраны. Интересно, что, несмотря на наличие клеточной стенки толщиной в несколько 100 нм, метод, производный от TIRF, известный как эпифлуоресцентная микроскопия с переменным углом (VAEM), недавно был разработан у растений для визуализации переноса везикул и событий слияния на плазматической мембране. В методике используется лазерный луч с неоптимальным углом падения, немного меньшим критического угла. Конопка и др. смогли визуализировать динамику мембранных белков, органелл и цитоскелета в различных растительных клетках (Konopka and Bednarek, 2008b; Wan et al., 2011) и подтвердили оптические принципы, лежащие в основе VAEM в растительных клетках. Они подробно проанализировали подвижность легких цепей клатрина (CLC) во время эндоцитоза (Konopka et al., 2008). Как в кончиках растущих корневых волосков, так и в диффузно разрастающихся эпидермальных клетках слитый белок CLC-XFP локализован вместе с родственными динамину белками DRP1C-GFP в динамических очагах в корковом слое клеток.Авторы смогли проследить появление и исчезновение белков на плазматической мембране и дифференцировать поведение двух различных DRP (Konopka and Bednarek, 2008a). Результаты демонстрируют участие DRP в клатрин-опосредованной мембранной динамике. Поскольку GFP-CESA3 можно визуализировать в живых клетках с помощью VAEM (Рис.
В методике используется лазерный луч с неоптимальным углом падения, немного меньшим критического угла. Конопка и др. смогли визуализировать динамику мембранных белков, органелл и цитоскелета в различных растительных клетках (Konopka and Bednarek, 2008b; Wan et al., 2011) и подтвердили оптические принципы, лежащие в основе VAEM в растительных клетках. Они подробно проанализировали подвижность легких цепей клатрина (CLC) во время эндоцитоза (Konopka et al., 2008). Как в кончиках растущих корневых волосков, так и в диффузно разрастающихся эпидермальных клетках слитый белок CLC-XFP локализован вместе с родственными динамину белками DRP1C-GFP в динамических очагах в корковом слое клеток.Авторы смогли проследить появление и исчезновение белков на плазматической мембране и дифференцировать поведение двух различных DRP (Konopka and Bednarek, 2008a). Результаты демонстрируют участие DRP в клатрин-опосредованной мембранной динамике. Поскольку GFP-CESA3 можно визуализировать в живых клетках с помощью VAEM (Рис. 1), будет интересно посмотреть, опосредована ли этим путем и интернализация CSCs. С помощью VAEM детально изучена организация и динамика актиновых филаментов в эпидермальных клетках гипокотилей Arabidopsis , а также показано постепенное появление переносчика оттока ауксина PIN2-GFP на наружной латеральной плазматической мембране корня. эпидермальные клетки (Staiger et al., 2009; Кляйне-Вен и др., 2011). Интересно, что метод VAEM можно использовать и для анализа олигомерного состояния белков. Количество субъединиц и стехиометрию мембраносвязанных белков можно определить с помощью статистического анализа стадий обесцвечивания GFP, слитого с белками. Ли и др. (2011) визуализировали меченый GFP аквапорин PIP2;1 в плазматической мембране корневых клеток. Генетическое присоединение GFP к PIP2;1 гарантировало, что каждый полипептид PIP2;1 был помечен только одним флуорофором.Чтобы определить олигомерное состояние PIP2;1, они подсчитали этапы фотообесцвечивания для выбранных флуоресцентных пятен.
1), будет интересно посмотреть, опосредована ли этим путем и интернализация CSCs. С помощью VAEM детально изучена организация и динамика актиновых филаментов в эпидермальных клетках гипокотилей Arabidopsis , а также показано постепенное появление переносчика оттока ауксина PIN2-GFP на наружной латеральной плазматической мембране корня. эпидермальные клетки (Staiger et al., 2009; Кляйне-Вен и др., 2011). Интересно, что метод VAEM можно использовать и для анализа олигомерного состояния белков. Количество субъединиц и стехиометрию мембраносвязанных белков можно определить с помощью статистического анализа стадий обесцвечивания GFP, слитого с белками. Ли и др. (2011) визуализировали меченый GFP аквапорин PIP2;1 в плазматической мембране корневых клеток. Генетическое присоединение GFP к PIP2;1 гарантировало, что каждый полипептид PIP2;1 был помечен только одним флуорофором.Чтобы определить олигомерное состояние PIP2;1, они подсчитали этапы фотообесцвечивания для выбранных флуоресцентных пятен. Максимальное количество стадий фотообесцвечивания элементарного GFP в индивидуальном токе было четыре, что согласуется с тетрамером аквапорина, наблюдаемым в кристаллографических исследованиях (Tornroth-Horsefield et al., 2006). Этот подход будет очень полезен для изучения стехиометрии биосинтетических комплексов клеточной стенки.
Максимальное количество стадий фотообесцвечивания элементарного GFP в индивидуальном токе было четыре, что согласуется с тетрамером аквапорина, наблюдаемым в кристаллографических исследованиях (Tornroth-Horsefield et al., 2006). Этот подход будет очень полезен для изучения стехиометрии биосинтетических комплексов клеточной стенки.
РИСУНОК 1. Визуализация GFP-CESA3 с использованием различных методов микроскопии. Выращенные в темноте клетки гипокотиля арабидопсиса , экспрессирующие каталитическую субъединицу целлюлозосинтазы CESA3, слитую с GFP, визуализировали с помощью конфокальной микроскопии с вращающимся диском (A) или эпифлуоресцентной микроскопии с переменным углом (B) . Обратите внимание, что в (B) хорошо видны только везикулы на поверхности клетки, и благодаря использованию объектива ×100 с числовой апертурой 1,49 разрешение xy и отношение сигнал/шум вблизи клетки края улучшаются.Шкала баров: 10 мкМ. Изображения из временного ряда (C – D) . Обратите внимание на появление везикулы GFP-CESA3 на втором изображении. Изображения из временного ряда (E–F) . Обратите внимание на исчезновение везикулы GFP-CESA3. Синие стрелки указывают на наличие или отсутствие GFP-CESA3 на последовательных изображениях.
Обратите внимание на появление везикулы GFP-CESA3 на втором изображении. Изображения из временного ряда (E–F) . Обратите внимание на исчезновение везикулы GFP-CESA3. Синие стрелки указывают на наличие или отсутствие GFP-CESA3 на последовательных изображениях.
В последнее время новые методы с высоким разрешением позволяют получать изображения за пределами барьера дифракционного разрешения. Используется несколько стратегий: микроскопия истощения стимулированного излучения (STED), микроскопия линейного или нелинейного структурированного освещения (SIM) и пуантилистские методы: фотоактивируемая локализационная микроскопия (PALM), флуоресцентная фотоактивируемая локализационная микроскопия (FPALM) и стохастическая оптическая реконструкция. микроскопия (ШТОРМ).Хотя они показывают большие перспективы для исследования клеточных стенок, все эти методы имеют свои сильные и слабые стороны. Подробное обсуждение см. в Leung and Chou (2011). Такие методы будут иметь большое значение для дальнейшего анализа организации CSCs и их связи с корковыми микротрубочками. STED уже использовался, чтобы показать, что переносчики оттока ауксинов PIN1 и PIN2 кластеризуются в микродоменах плазматической мембраны (Kleine-Vehn et al., 2011).
STED уже использовался, чтобы показать, что переносчики оттока ауксинов PIN1 и PIN2 кластеризуются в микродоменах плазматической мембраны (Kleine-Vehn et al., 2011).
Визуализация роста растений в 4D
Основными препятствиями для получения изображений 4D (3D + время) являются неотъемлемое ограничение скорости методов точечного сканирования для оптических срезов и фотообесцвечивание, сопровождающее многократное возбуждение.Использование световой флуоресцентной микроскопии (LSFM), известной в различных реализациях как селективное плоское освещение (SPIM) или цифровая сканирующая лазерная световая микроскопия (DSLM), преодолевает эти ограничения (Huisken et al., 2004; Keller and Stelzer). , 2008). Этот метод основан на использовании тонкого листа света, который освещает одну плоскость X-Y образца, в сочетании с широкопольным флуоресцентным детектированием с помощью ПЗС-камеры в направлении, перпендикулярном плоскости возбуждения.При этом методе освещается только узкая область вблизи плоскости наблюдения, в результате чего флуорофоры за пределами этой плоскости не возбуждаются, что снижает фоновые сигналы и фотоповреждение. Эти инструментальные характеристики создают эффект оптического сечения, улучшают контраст и позволяют проводить длительные наблюдения за чувствительными образцами. Трехмерная запись осуществляется путем перемещения образца через освещаемую плоскость. Получение серии изображений под тремя разными углами в сочетании с алгоритмом обработки позволяет отличить флуоресценцию сигнала от рассеянного фонового света.Этот метод недавно был использован для регистрации роста вертикально ориентированных кончиков первичных корней и зачатков боковых корней в течение до 38 часов без каких-либо явных повреждений (Maizel et al., 2011). Кроме того, можно было визуализировать динамику эндосомальных компартментов в течение более 40 с со скоростью 4 кадра в секунду. Этот метод позволит делать 3D-реконструкции целых органов и отслеживать рост каждой отдельной клетки в течение длительных периодов времени.
Эти инструментальные характеристики создают эффект оптического сечения, улучшают контраст и позволяют проводить длительные наблюдения за чувствительными образцами. Трехмерная запись осуществляется путем перемещения образца через освещаемую плоскость. Получение серии изображений под тремя разными углами в сочетании с алгоритмом обработки позволяет отличить флуоресценцию сигнала от рассеянного фонового света.Этот метод недавно был использован для регистрации роста вертикально ориентированных кончиков первичных корней и зачатков боковых корней в течение до 38 часов без каких-либо явных повреждений (Maizel et al., 2011). Кроме того, можно было визуализировать динамику эндосомальных компартментов в течение более 40 с со скоростью 4 кадра в секунду. Этот метод позволит делать 3D-реконструкции целых органов и отслеживать рост каждой отдельной клетки в течение длительных периодов времени.
Заключение
Теперь доступен большой набор методов флуоресцентной визуализации для изучения тканей, клеток и клеточных компонентов в 3D или 4D у растений. Эти методы визуализации, скорее всего, произведут революцию в ближайшие годы в нашем понимании динамики архитектуры клеточной стенки и клеточных процессов, участвующих в синтезе компонентов клеточной стенки. Однако каждый метод визуализации имеет относительные сильные и слабые стороны, которые необходимо учитывать при решении конкретной биологической проблемы.
Эти методы визуализации, скорее всего, произведут революцию в ближайшие годы в нашем понимании динамики архитектуры клеточной стенки и клеточных процессов, участвующих в синтезе компонентов клеточной стенки. Однако каждый метод визуализации имеет относительные сильные и слабые стороны, которые необходимо учитывать при решении конкретной биологической проблемы.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Подтверждение
Авторы благодарят Джима Домпьера (Plateforme intégrée Imagif, Gif S/Yvette) за помощь в экспериментах с VAEM.
Сноски
Ссылки
Андерсон, К. Т., Кэрролл, А., Ахметова, Л., и Сомервилл, К. (2010). Визуализация в реальном времени переориентации целлюлозы во время расширения клеточной стенки в корнях Arabidopsis . Завод физиол. 152, 787–796.
152, 787–796.
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Андерсон, К.Т., Уоллес И.С. и Сомервилль Ч.Р. (2012). Метаболическая маркировка с помощью аналога фукозы выявляет доставку, архитектуру и динамику пектина в клеточных стенках Arabidopsis . Проц. Натл. акад. науч. США 109, 1329–1334.
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Бишофф, В., Деспре, Т., Муйе, Г., Вернет, С., Гонно, М., и Хофте, Х. (2011). Фитохромная регуляция синтеза целлюлозы у арабидопсиса . Курс. биол. 21, 1822–1827 гг.
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Брача-Дрори, К., Шихрур, К., Кац, А., Олива, М., Ангеловичи, Р., Яловски, С., и Охад, Н. (2004). Обнаружение белок-белковых взаимодействий в растениях с помощью бимолекулярной флуоресцентной комплементации. Завод J . 40, 419–427.
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Брингманн М. , Ли Э., Сампаткумар А., Коджабек Т., Хаузер, М.Т., и Перссон, С. (2012). Взаимодействие POM-POM2/целлюлозосинтазы1 необходимо для функциональной ассоциации целлюлозосинтазы и микротрубочек в Arabidopsis . Растительная клетка 24, 163–177.
, Ли Э., Сампаткумар А., Коджабек Т., Хаузер, М.Т., и Перссон, С. (2012). Взаимодействие POM-POM2/целлюлозосинтазы1 необходимо для функциональной ассоциации целлюлозосинтазы и микротрубочек в Arabidopsis . Растительная клетка 24, 163–177.
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Чанг, П. В., Прешер, Дж. А., Слеттен, Э. М., Баскин, Дж. М., Миллер, И. А., Агард, Н. Дж., Ло, А., и Бертоцци, К. Р. (2010). Щелчковая химия без меди у живых животных. Проц.Натл. акад. науч. США 107, 1821–1826 гг.
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Чен, С., Эрхардт, Д. В., и Сомервилл, Ч. Р. (2010). Мутации сайтов фосфорилирования синтазы целлюлозы (CESA1) модулируют анизотропную экспансию клеток и двунаправленную подвижность синтазы целлюлозы. Проц. Натл. акад. науч. США 107, 17188–17193.
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Кроуэлл, Э. Ф., Бишофф, В., Desprez, T., Rolland, A., Stierhof, YD, Schumacher, K., Gonneau, M., Hofte, H., и Vernhettes, S. (2009). Приостановка телец Гольджи на микротрубочках регулирует секрецию комплексов целлюлозосинтазы у Arabidopsis . Растительная клетка 21, 1141–1154.
Ф., Бишофф, В., Desprez, T., Rolland, A., Stierhof, YD, Schumacher, K., Gonneau, M., Hofte, H., и Vernhettes, S. (2009). Приостановка телец Гольджи на микротрубочках регулирует секрецию комплексов целлюлозосинтазы у Arabidopsis . Растительная клетка 21, 1141–1154.
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
ДеБолт, С., Гутьеррес, Р., Эрхардт, Д. В., Мело, К. В., Росс, Л., Катлер, С. Р., Сомервиль, К., и Бонетта, Д. (2007a). Морлин, ингибитор динамики кортикальных микротрубочек и движения целлюлозосинтазы. Проц. Натл. акад. науч. США 104, 5854–5859.
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
ДеБолт, С., Гутьеррес, Р., Эрхардт, Д. В., и Сомервиль, К. (2007b). Неподвижные субъединицы синтазы целлюлозы неоднократно накапливаются в локализованных областях плазматической мембраны в клетках гипокотиля Arabidopsis после обработки 2,6-дихлорбензонитрилом. Завод Физиол . 145, 334–338.
Завод Физиол . 145, 334–338.
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Депре, Т., Juraniec, M., Crowell, E.F., Jouy, H., Pochylova, Z., Parcy, F., Hofte, H., Gonneau, M., и Vernhettes, S. (2007). Организация комплексов целлюлозосинтазы, участвующих в синтезе первичной клеточной стенки у Arabidopsis thaliana . Проц. Натл. акад. науч. США 104, 15572–15577.
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Дхонукше, П., Аниенто, Ф., Хван, И., Робинсон, Д.Г., Мравец, Дж., Штирхоф, Ю.Д., и Фримл, Дж. (2007). Опосредованный клатрином конститутивный эндоцитоз носителей оттока PIN-ауксина у Arabidopsis . Курс. биол. 17, 520–527.
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Домозич, Д. С., Ламбьяс, Л., Кимле, С. Н., и Гретц, М. Р. (2009). Развитие клеточной стенки и биполярный рост Desmid Penium margaritaceum (Zygnematophyceae, Streptophyta). Асимметрия в симметричном мире. J. Phycol. 45, 879–893.
Асимметрия в симметричном мире. J. Phycol. 45, 879–893.
Полнотекстовая перекрестная ссылка
Граф Р., Ритдорф Дж. и Циммерманн Т. (2005). Микроскопия живых клеток с вращающимся диском. Доп. Биохим. англ. Биотехнолог. 95, 57–75.
Опубликовано Резюме | Опубликован полный текст
Гу, Ю., Каплински, Н., Брингманн, М., Кобб, А., Кэрролл, А., Сампаткумар, А., Баскин, Т., Перссон, С., и Сомервилл, К. (2010). Идентификация нового CESA-ассоциированного белка, необходимого для биосинтеза целлюлозы. Проц. Натл. акад. науч. США 107, 12866–12871.
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Гутьеррес, Р., Линдебум, Дж.Дж., Паредес А.Р., Эмонс А.М. и Эрхардт Д.В. (2009). Кортикальные микротрубочки Arabidopsis направляют доставку синтазы целлюлозы к плазматической мембране и взаимодействуют с транспортными отсеками синтазы целлюлозы. Нац. Клеточная биол. 11, 797–806.
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Huisken, J., Swoger, J., Del Bene, F., Wittbrodt, J., and Stelzer, E.H. (2004). Оптические срезы глубоко внутри живых эмбрионов с помощью микроскопии с селективным плоским освещением. Наука 305, 1007–1009.
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Ибатуллин Ф.М., Банасиак А., Бауманн М.Дж., Греффе Л., Такахаши Дж., Меллерович Э.Дж. и Брумер Х. (2009). Флуорогенный анализ в реальном времени для визуализации активности гликозидгидролазы в растениях. Завод физиол. 151, 1741–1750.
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Келлер, П. Дж., и Стельцер, Э. Х.(2008). Количественная визуализация целых эмбрионов in vivo с помощью цифровой флуоресцентной микроскопии с лазерным сканированием. Курс. мнение Нейробиол. 18, 624–632.
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Кляйне-Вен, Дж. , Вабник, К., Мартиньер, А., Ланговски, Л., Уиллиг, К., Нарамото, С., Лейтнер, Дж., Танака, Х., Якобс, С., Роберт, С., Люшниг К., Говертс В., Хелл С.В., Рунионс Дж. и Фримл Дж. (2011). Рециркуляция, кластеризация и эндоцитоз совместно поддерживают полярность переносчика ауксина PIN на плазматической мембране. Мол. Сист. биол. 7, 540.
, Вабник, К., Мартиньер, А., Ланговски, Л., Уиллиг, К., Нарамото, С., Лейтнер, Дж., Танака, Х., Якобс, С., Роберт, С., Люшниг К., Говертс В., Хелл С.В., Рунионс Дж. и Фримл Дж. (2011). Рециркуляция, кластеризация и эндоцитоз совместно поддерживают полярность переносчика ауксина PIN на плазматической мембране. Мол. Сист. биол. 7, 540.
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Конопка, К.А., Бэкьюс, С.К., и Беднарек, С.Ю. (2008). Динамика родственного динамину белка 1C Arabidopsis и легкой цепи клатрина на плазматической мембране. Растительная клетка 20, 1363–1380.
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Конопка, К.А., и Беднарек, С.Ю. (2008a). Сравнение динамики и функциональной избыточности родственных динамину изоформ DRP1A и DRP1C Arabidopsis во время развития растений. Завод Физиол . 147, 1590–1602 гг.
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Li, X. , Wang, X., Yang, Y., Li, R., He, Q., Fang, X., Luu, D.T., Maurel, C., and Lin, J. (2011). Одномолекулярный анализ динамики и разделения PIP2; 1 выявляет множественные способы регуляции аквапоринов плазматической мембраны Arabidopsis . Растительная клетка 23, 3780–3797.
, Wang, X., Yang, Y., Li, R., He, Q., Fang, X., Luu, D.T., Maurel, C., and Lin, J. (2011). Одномолекулярный анализ динамики и разделения PIP2; 1 выявляет множественные способы регуляции аквапоринов плазматической мембраны Arabidopsis . Растительная клетка 23, 3780–3797.
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Майзель, А., фон Вангенхайм, Д., Федеричи, Ф., Хазелофф, Дж., и Стельцер, Э. Х. (2011). Живая визуализация роста растений с высоким разрешением при ярком освещении, близком к физиологическому, с использованием световой флуоресцентной микроскопии. Завод J . 68, 377–385.
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Маркус, С.Э., Блейк, А.В., Бенианс, Т.А., Ли, К.Дж., Пойсер, К., Дональдсон, Л., Леру, О., Роговски, А., Петерсен, Х.Л., Борастон, А., Гилберт, Х.Дж., Уиллатс, В.Г., и Нокс, Дж.П. (2010). Ограниченный доступ белков к маннановым полисахаридам в интактных клеточных стенках растений. Завод J. 64, 191–203.
Завод J. 64, 191–203.
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Маркус, С.Э., Верхертбругген, Ю., Эрве, К., Ордаз-Ортис, Дж.Дж., Фаркас, В., Педерсен, Х.Л., Уиллатс, В.Г., и Нокс, Дж.П. (2008). Пектиновый гомогалактуронан маскирует обильные наборы ксилоглюкановых эпитопов в стенках клеток растений. BMC Растение Биол. 8, 60. doi: 10.1186/1471-2229-8-60
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Моллер, И., Маркус, С.Э., Хаегер, А., Верхертбругген, Ю., Верхоф, Р., Шолс, Х., Ульвсков, П., Миккельсен, Дж.Д., Нокс, Дж.П., и Уиллатс, В. (2008). Высокопроизводительный скрининг моноклональных антител против гликанов клеточной стенки растений путем иерархической группировки их профилей связывания углеводного микрочипа. Гликоконж. Дж. 25, 37–48.
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Нишикубо, Н. , Такахаши, Дж., Роос, А. А., Дерба-Макелуч, М., Пиенс, К., Брумер, Х., Teeri, T.T., Stalbrand, H., and Mellerowicz, EJ (2011). Перестройки ксилоглюкана, опосредованные эндо-трансгликозилазой, в развивающейся древесине гибридной осины. Завод физиол. 155, 399–413.
, Такахаши, Дж., Роос, А. А., Дерба-Макелуч, М., Пиенс, К., Брумер, Х., Teeri, T.T., Stalbrand, H., and Mellerowicz, EJ (2011). Перестройки ксилоглюкана, опосредованные эндо-трансгликозилазой, в развивающейся древесине гибридной осины. Завод физиол. 155, 399–413.
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Паттатил, С., Авчи, У., Болдуин, Д., Свеннес, А.Г., Макгилл, Дж.А., Поппер, З., Буттен, Т., Альберт, А., Дэвис, Р.Х., Ченнаредди, К., Донг, Р., О’Ши Б., Росси Р., Леофф К., Фрешур Г., Нарра Р., О’Нил М., York, WS, and Hahn, MG (2010). Комплексный набор моноклональных антител к гликанам клеточной стенки растений. Завод Физиол . 153, 514–525.
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Ralet, M.C., Tranquet, O., Poulain, D., Moise, A., and Guillon, F. (2010). Моноклональные антитела к рамногалактуронановому остову I. Планта 231, 1373–1383.
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Рулофсен, П. А. и Хаувинк, А.Л. (1953). Архитектура и рост первичной клеточной стенки в волосках некоторых растений и спорангиофоре Phycomyces. Акта Бот. Нирланд 2, 218–225.
А. и Хаувинк, А.Л. (1953). Архитектура и рост первичной клеточной стенки в волосках некоторых растений и спорангиофоре Phycomyces. Акта Бот. Нирланд 2, 218–225.
Санчес-Родригес, К., Бауэр, С., Хемати, К., Сакс, Ф., Ибанез, А.Б., Водермайер, В., Конлехнер, К., Сампаткумар, А., Руггеберг, М., Айхингер, Э. ., Нойметцлер, Л., Бургерт, И., Сомервилль, К., Хаузер, М.Т., и Перссон, С. (2012). Chitinase-like1/pom-pom1 и его гомолог CTL2 представляют собой глюкан-взаимодействующие белки, важные для биосинтеза целлюлозы в Arabidopsis . Растительная клетка 24, 589–607.
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Сава, М., Хсу, Т.Л., Ито, Т., Сугияма, М., Хэнсон, С.Р., Фогт, П.К., и Вонг, С.Х. (2006). Гликопротеомные зонды для флуоресцентной визуализации фукозилированных гликанов in vivo. Проц. Натл. акад. науч. США 103, 12371–12376.
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Сориано Дель Амо, Д. , Ван, В., Цзян, Х., Безансени, К., Yan, A.C., Levy, M., Liu, Y., Marlow, F.L., and Wu, P. (2010). Биосовместимые катализаторы на основе меди (I) для визуализации гликанов in vivo. Дж. Ам. хим. соц. 132, 16893–16899.
, Ван, В., Цзян, Х., Безансени, К., Yan, A.C., Levy, M., Liu, Y., Marlow, F.L., and Wu, P. (2010). Биосовместимые катализаторы на основе меди (I) для визуализации гликанов in vivo. Дж. Ам. хим. соц. 132, 16893–16899.
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Staiger, C.J., Sheahan, M.B., Khurana, P., Wang, X., McCurdy, D.W., and Blanchoin, L. (2009). В динамике актиновых филаментов преобладают быстрый рост и активность в корковом массиве Arabidopsis . J. Cell Biol. 184, 269–280.
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Саттер, Дж. У., Кампанони, П., Тиррелл, М., и Блатт, М. Р. (2006). Селективную подвижность и чувствительность к SNARE демонстрирует канал Arabidopsis KAT1 K + на плазматической мембране. Растительная клетка 18, 935–954.
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Такахаши Дж. , Рудсандер У. Дж., Хеденстрем М., Банасиак А., Хархольт, Дж., Амелот, Н., Иммерзил, П., Райден, П., Эндо, С., Ибатуллин, Ф.М., Брумер, Х., дель Кампильо, Э., Мастер, Э.Р., Шеллер, Х.В., Сандберг , Б., Тири, Т.Т., и Меллерович, Э.Дж. (2009). KORRIGAN1 и его гомолог осины PttCel9A1 снижают кристалличность целлюлозы в стеблях Arabidopsis . Физиол растительных клеток . 50, 1099–1115.
, Рудсандер У. Дж., Хеденстрем М., Банасиак А., Хархольт, Дж., Амелот, Н., Иммерзил, П., Райден, П., Эндо, С., Ибатуллин, Ф.М., Брумер, Х., дель Кампильо, Э., Мастер, Э.Р., Шеллер, Х.В., Сандберг , Б., Тири, Т.Т., и Меллерович, Э.Дж. (2009). KORRIGAN1 и его гомолог осины PttCel9A1 снижают кристалличность целлюлозы в стеблях Arabidopsis . Физиол растительных клеток . 50, 1099–1115.
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Timmers, J., Vernhettes, S., Desprez, T., Vincken, J.P., Visser, R.G., and Trindade, LM (2009). Взаимодействия между мембраносвязанными синтазами целлюлозы, участвующими в синтезе вторичной клеточной стенки. ФЭБС Письмо. 583, 978–982.
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Tornroth-Horsefield, S., Wang, Y., Hedfalk, K., Johanson, U., Karlsson, M., Tajkhorshid, E., Neutze, R., and Kjellbom, P. (2006). Структурный механизм аквапоринового гейтирования растений. Природа 439, 688–694.
Природа 439, 688–694.
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Виссенберг, К., Фрай, С.К., Поли, М., Хофте, Х., и Вербелен, Дж.П. (2005). XTH действует на поверхности раздела микрофибрилл-матрицы во время удлинения клеток. Дж. Экспл. Бот. 56, 673–683.
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Ван, Ю., Эш, В. М. III., Фань, Л., Хао, Х., Ким, М. К., и Лин, Дж. (2011). Флуоресцентная микроскопия полного внутреннего отражения с переменным углом интактных клеток Arabidopsis thaliana . Растительные методы 7, 27.
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Вайтман, Р., Маршалл, Р., и Тернер, С.Р. (2009). Компартмент, содержащий синтазу целлюлозы, быстро перемещается под участками синтеза вторичной стенки. Физиол растительных клеток . 50, 584–594.
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки
Анализ экспрессированных меток последовательности и идентификация генов, кодирующих ферменты, разрушающие клеточную стенку, грибоядной нематоды Aphelenchus avenae | BMC Genomics
Генерация EST из библиотеки кДНК
A. avenae
avenae A смешанная стадия A.Библиотека кДНК avenae (Aamk) была сконструирована для получения EST (таблица 1). Шестнадцать клонов были выбраны случайным образом, и размеры вставок в этих клонах были оценены после расщепления соответствующими ферментами рестрикции. Размеры этих вставок варьировались от 400 до 1600 пар оснований (п.н.), в среднем 1,1 тыс. пар оснований (т.п.н.). В общей сложности 5472 клона кДНК были впоследствии случайным образом выделены и секвенированы с 5′-конца для получения EST. Последовательности были удалены от векторной последовательности, адаптерной последовательности, поли(А)-хвоста и последовательности низкого качества и отфильтрованы по минимальной длине (150 п.н.), в результате чего было получено в общей сложности 5076 EST высокого качества.Средняя длина представленных EST составила 468 нуклеотидов (нт).
Таблица 1 A. avenae Библиотека кДНК, статистика EST и кластеровФормирование и анализ кластеров
Для выявления перекрывающихся последовательностей EST, повышения точности оснований и длины транскриптов, а также для получения неповторяющихся данных EST для дальнейшей функциональной аннотации и сравнительного анализа, 5076 EST из библиотеки A. avenae были сгруппированы по идентичности последовательностей в кластеры. На основе областей идентичности нуклеотидов последовательности EST были объединены в смежные консенсусные последовательности (контиги).EST-члены «контигов» происходят из идентичных транскриптов, в то время как члены «кластеров» могут происходить из одного и того же гена, но представлять разные изоформы сплайсинга транскриптов (, т.е. EST образуют контиги, контиги образуют кластеры). Из EST были сгенерированы две тысячи семьсот неизбыточных кластеров EST (таблица 1). В 2005 случаях кластеры состоят из одного EST, тогда как самый большой отдельный кластер содержит 81 EST (1 случай) (рис. 2). Большинство контигов (650 из 695) состояло из 2-10 EST.Было обнаружено, что 89 кластеров содержат несколько членов контигов, что указывает на потенциальные изоформы сплайсинга. Благодаря устранению избыточности во время построения этого контига общее количество нуклеотидов, используемых для дальнейшего анализа, было уменьшено с 2,82 миллиона до 1,61 миллиона.
avenae были сгруппированы по идентичности последовательностей в кластеры. На основе областей идентичности нуклеотидов последовательности EST были объединены в смежные консенсусные последовательности (контиги).EST-члены «контигов» происходят из идентичных транскриптов, в то время как члены «кластеров» могут происходить из одного и того же гена, но представлять разные изоформы сплайсинга транскриптов (, т.е. EST образуют контиги, контиги образуют кластеры). Из EST были сгенерированы две тысячи семьсот неизбыточных кластеров EST (таблица 1). В 2005 случаях кластеры состоят из одного EST, тогда как самый большой отдельный кластер содержит 81 EST (1 случай) (рис. 2). Большинство контигов (650 из 695) состояло из 2-10 EST.Было обнаружено, что 89 кластеров содержат несколько членов контигов, что указывает на потенциальные изоформы сплайсинга. Благодаря устранению избыточности во время построения этого контига общее количество нуклеотидов, используемых для дальнейшего анализа, было уменьшено с 2,82 миллиона до 1,61 миллиона. Кроме того, этот процесс значительно увеличил длину собранных последовательностей транскриптов с 468 ± 114 нуклеотидов для представленных EST только до 595 ± 321 нуклеотидов для контигов. Самая длинная сгенерированная последовательность также увеличилась с 724 до 2154 н.
Кроме того, этот процесс значительно увеличил длину собранных последовательностей транскриптов с 468 ± 114 нуклеотидов для представленных EST только до 595 ± 321 нуклеотидов для контигов. Самая длинная сгенерированная последовательность также увеличилась с 724 до 2154 н.
Гистограмма, показывающая распределение EST от A.avenae по размеру кластера . Например, было пять кластеров размером 23, содержащих в общей сложности 115 EST. Распределение размеров контигов не показано.
На основе идентифицированных кластеров было идентифицировано 2700 генов A. avenae , что соответствует коэффициенту обнаружения новых генов 53% (2700/5076). Однако 2700 кластеров, вероятно, будут завышенной оценкой истинной скорости обнаружения генов, поскольку один ген может быть представлен несколькими неперекрывающимися кластерами.Такая «фрагментация» оценивается в 18% при использовании C. elegans в качестве эталонного генома [39]. После учета такой потенциальной фрагментации мы подсчитали, что последовательностей A. avenae произошли от минимум 2214 генов, что дает степень обнаружения 44% (2214/5076). Предполагая, что общее количество генов составляет от 14 000 до 21 000, диапазон охватывает Meloidogyne hapla [40], M. incognita [41] и C. elegans (Wormpep v. 203), набор кластерных данных может представлять примерно 11-16 % от А.avenae генов.
После учета такой потенциальной фрагментации мы подсчитали, что последовательностей A. avenae произошли от минимум 2214 генов, что дает степень обнаружения 44% (2214/5076). Предполагая, что общее количество генов составляет от 14 000 до 21 000, диапазон охватывает Meloidogyne hapla [40], M. incognita [41] и C. elegans (Wormpep v. 203), набор кластерных данных может представлять примерно 11-16 % от А.avenae генов.
Обилие транскриптов и широко представленные гены
Высокий уровень представленности в библиотеке кДНК обычно коррелирует с высоким обилием транскриптов в исходном биологическом образце [42], хотя артефакты конструирования библиотеки могут привести к отбору за или против некоторых транскриптов. Кластеры A. avenae были ранжированы в соответствии с количеством участвующих EST, и 25 лучших кластеров приведены в таблице 2. Каждый из этих кластеров содержал пятнадцать или более копий EST и составлял 16% от общего числа полученных EST. Восемнадцать из кластеров имели значительные совпадения с генами с аннотированными функциями на основе BLASTX (E < 1e-5) по сравнению с неизбыточной базой данных, и все они имели гомологи у нематод. Транскрипты, широко представленные в библиотеке A. avenae , включали гены, кодирующие структурные белки (такие как актин, коллаген, тропомиозин и тропонин С) и белки, которые осуществляют основные метаболические процессы (, например, . цитохром с-оксидаза, АТФ-синтаза). Другие распространенные EST включали небольшой белок теплового шока и фосфоенолпируваткарбоксикиназу.Последний фермент ранее был клонирован из паразитических нематод Haemonchus contortus и Ascaris suum [43, 44]. Кластер AAC00541, содержащий 23 EST, был подобен белку семейства SXP/RAL-2 паразитической нематоды Anisakis simplex . Подобные гены ранее были охарактеризованы у нематод, паразитирующих на растениях [45], и было показано, что отдельные гены экспрессируются в ряде секреторных тканей, включая клетки желез, окружающие основные органы чувств (амфиды) и гиподерму.
Восемнадцать из кластеров имели значительные совпадения с генами с аннотированными функциями на основе BLASTX (E < 1e-5) по сравнению с неизбыточной базой данных, и все они имели гомологи у нематод. Транскрипты, широко представленные в библиотеке A. avenae , включали гены, кодирующие структурные белки (такие как актин, коллаген, тропомиозин и тропонин С) и белки, которые осуществляют основные метаболические процессы (, например, . цитохром с-оксидаза, АТФ-синтаза). Другие распространенные EST включали небольшой белок теплового шока и фосфоенолпируваткарбоксикиназу.Последний фермент ранее был клонирован из паразитических нематод Haemonchus contortus и Ascaris suum [43, 44]. Кластер AAC00541, содержащий 23 EST, был подобен белку семейства SXP/RAL-2 паразитической нематоды Anisakis simplex . Подобные гены ранее были охарактеризованы у нематод, паразитирующих на растениях [45], и было показано, что отдельные гены экспрессируются в ряде секреторных тканей, включая клетки желез, окружающие основные органы чувств (амфиды) и гиподерму.
Семь из 25 наиболее широко представленных транскриптов из A. avenae не имели значительного сходства с какой-либо последовательностью в базе данных неизбыточных белков (таблица 2). ). Поскольку большинство данных о нематодах доступны только в виде EST и, следовательно, не включены в анализ BLASTX, мы сравнили эти 7 контигов с dbEST, используя BLASTN и TBLASTX. Однако эти поиски не дали значимых совпадений (E < 1e-5).Мы также провели поиск BLASTN и TBLASTX в базе данных неизбыточных нуклеотидов для этих последовательностей. Шесть кластеров не дали никаких совпадений из этой базы данных, но кластер AAC00148 дал совпадение с использованием анализа TBLASTX (E < 1e-5) (таблица 2).
Сравнения с белками других видов
Мы сравнили 2700 кластерных последовательностей из A. avenae с тремя базами данных, содержащими белковые последовательности разных организмов. Последовательности кластеров сравнивали с последовательностями белков из (i) C.elegans (WORMPEP v.203 [46]), (ii) другие нематоды (доступные белковые последовательности и пептиды из концептуально транслируемых EST) и (iii) организмы, отличные от нематод (из базы данных неизбыточных белков NCBI) [47] . 66% из кластеров A. avenae (1782 из 2700) имели совпадения в одной или нескольких из трех баз данных, и эти совпадения были представлены с помощью SimiTri [48] (рис. 3). В большинстве случаев обнаружения гомологий (1242/1782) совпадения были обнаружены во всех трех обследованных базах данных.Генные продукты в этой категории, как правило, широко распространены среди многоклеточных животных, и многие из них участвуют в основных биологических процессах. Изучение отдельных поисковых запросов в базе данных показало, что 1567 (58,0%) имели гомологи в C. elegans , 1750 (64,8%) в других нематодах и 1321 (48,9%) в организмах, отличных от нематод. 918 кластеров (34,0%), которые не имели значительного сходства с какими-либо последовательностями в этих трех базах данных белков, искали в базах данных неизбыточных нуклеотидов и dbEST с использованием BLASTN и TBLASTX (используя отсечку 1e-05).
Последовательности кластеров сравнивали с последовательностями белков из (i) C.elegans (WORMPEP v.203 [46]), (ii) другие нематоды (доступные белковые последовательности и пептиды из концептуально транслируемых EST) и (iii) организмы, отличные от нематод (из базы данных неизбыточных белков NCBI) [47] . 66% из кластеров A. avenae (1782 из 2700) имели совпадения в одной или нескольких из трех баз данных, и эти совпадения были представлены с помощью SimiTri [48] (рис. 3). В большинстве случаев обнаружения гомологий (1242/1782) совпадения были обнаружены во всех трех обследованных базах данных.Генные продукты в этой категории, как правило, широко распространены среди многоклеточных животных, и многие из них участвуют в основных биологических процессах. Изучение отдельных поисковых запросов в базе данных показало, что 1567 (58,0%) имели гомологи в C. elegans , 1750 (64,8%) в других нематодах и 1321 (48,9%) в организмах, отличных от нематод. 918 кластеров (34,0%), которые не имели значительного сходства с какими-либо последовательностями в этих трех базах данных белков, искали в базах данных неизбыточных нуклеотидов и dbEST с использованием BLASTN и TBLASTX (используя отсечку 1e-05). 56 кластеров генерировали совпадения в этих поисках, но для остальных 862 последовательностей (31,9 %) совпадений не было получено.
56 кластеров генерировали совпадения в этих поисках, но для остальных 862 последовательностей (31,9 %) совпадений не было получено.
Сравнение A. Avenae A. Avenae Кластерные последовательности с
C. Elegans 3 , другие нематоды и базы данных белков белков нематода, использующие SIMITRI . Числа в каждой вершине указывают количество кластерных последовательностей, соответствующих только этой конкретной базе данных. Числа на ребрах указывают количество последовательностей кластеров, соответствующих двум базам данных, связанным этим ребром.Число внутри треугольника указывает количество генов A. avenae , совпадающих с последовательностями во всех трех базах данных. В таблице 3 показаны 15 генных продуктов с наивысшим уровнем консервативности (значение E в диапазоне от 0 до e-151) между A. avenae и C. elegans ; к ним относятся генные продукты, участвующие в клеточной структуре (например, актин, UNC-87), биосинтезе или регуляции белка (например, UBQ-1, фактор элонгации) и метаболизме (например, енолаза, цитохромоксидаза). Представленность этих кластеров в коллекции A. avenae EST варьировала от 81 до 2 EST. Ни один из этих наиболее консервативных генных продуктов не был специфичен в отношении нематод. Из всех кластеров 461 (17,1%) имели гомологию только с нематодами, либо C. elegans (4), либо с другими нематодами (137), либо с теми и другими (320) (рис. 3). Наиболее консервативным (1e-111) из этих специфичных для нематод белков был гомолог ингибитора сериновой протеиназы, ранее охарактеризованный у C. elegans (K10D3.4) и паразитические нематоды [49, 50]. Среди других наиболее консервативных кластеров, специфичных для нематод, были гомологи ранее охарактеризованных структурных белков C. elegans (например, кластер, AAC01973, соответствующий белку семейства коллагенов, COL-176), а также неохарактеризованные гипотетические белки C. elegans ( например, кластер AAC01948 соответствует гену C. elegans , C34E7.4, функция которого неизвестна).
Представленность этих кластеров в коллекции A. avenae EST варьировала от 81 до 2 EST. Ни один из этих наиболее консервативных генных продуктов не был специфичен в отношении нематод. Из всех кластеров 461 (17,1%) имели гомологию только с нематодами, либо C. elegans (4), либо с другими нематодами (137), либо с теми и другими (320) (рис. 3). Наиболее консервативным (1e-111) из этих специфичных для нематод белков был гомолог ингибитора сериновой протеиназы, ранее охарактеризованный у C. elegans (K10D3.4) и паразитические нематоды [49, 50]. Среди других наиболее консервативных кластеров, специфичных для нематод, были гомологи ранее охарактеризованных структурных белков C. elegans (например, кластер, AAC01973, соответствующий белку семейства коллагенов, COL-176), а также неохарактеризованные гипотетические белки C. elegans ( например, кластер AAC01948 соответствует гену C. elegans , C34E7.4, функция которого неизвестна).
 avenae и C. elegans
avenae и C. elegans 137 кластерных последовательностей, в которых гомологи присутствовали только у других нематод, были дополнительно классифицированы на основе их результатов BLAST (BLASTX и TBLASTX) (дополнительный файл 1). Совпадения были обнаружены у нематод, паразитирующих на растениях, животных и свободноживущих. 24,8% последовательностей (34 из 137) имели гомологию только с последовательностями фитопаразитических нематод. Некоторые из этих последовательностей были похожи на ранее охарактеризованные ферменты, разрушающие клеточную стенку, которые, как известно, участвуют в процессе паразитизма этих нематод.Например, кластер AAC01592 соответствовал экспансиноподобному белку из B. xylophilus [19], а кластер AAC02968 соответствовал предшественнику β-1,4-эндоглюканазы из Globodera rostochiensis [20]. Дальнейший анализ некоторых ферментов, разрушающих клеточную стенку, присутствующих в A. avenae , представлен ниже.
Идентификация транскриптов, сходных с генами реакции на стресс, связанными с высыханием
Поиски BLASTX (E < 1e-5) кластерных последовательностей A. avenae в базах данных nr белков позволили идентифицировать гены, которые могут кодировать белки или ферменты, важные для обеспечения защита от высыхания или другого стресса окружающей среды (Дополнительный файл 2).Одним примечательным наблюдением было присутствие последовательностей, сходных с белками позднего эмбрионального обилия (LEA), которые, как известно, связаны с толерантностью к водному стрессу в результате высыхания [9]. Агрегация белков во время высушивания, вероятно, представляет серьезную потенциальную опасность для ангидробиотов; Белки LEA могут действовать как молекулярные шапероны или молекулярные щиты и играть важную роль в предотвращении этой агрегации [51]. Тринадцать EST, распределенных по трем кластерам (AAC00729, AAC00888 и AAC01781), были идентифицированы как имеющие значительное сходство с белками LEA.Кластер, AAC01781, который был идентифицирован как синглтон, соответствовал ранее охарактеризованному белку LEA из высушенного A. avenae [9]. Кроме того, мы также идентифицировали множественные копии цитохрома Р450, супероксиддисмутазы, глутатионпероксидазы и глутатион-S-трансферазы, ферментов, участвующих в защите от окислительного повреждения.
avenae в базах данных nr белков позволили идентифицировать гены, которые могут кодировать белки или ферменты, важные для обеспечения защита от высыхания или другого стресса окружающей среды (Дополнительный файл 2).Одним примечательным наблюдением было присутствие последовательностей, сходных с белками позднего эмбрионального обилия (LEA), которые, как известно, связаны с толерантностью к водному стрессу в результате высыхания [9]. Агрегация белков во время высушивания, вероятно, представляет серьезную потенциальную опасность для ангидробиотов; Белки LEA могут действовать как молекулярные шапероны или молекулярные щиты и играть важную роль в предотвращении этой агрегации [51]. Тринадцать EST, распределенных по трем кластерам (AAC00729, AAC00888 и AAC01781), были идентифицированы как имеющие значительное сходство с белками LEA.Кластер, AAC01781, который был идентифицирован как синглтон, соответствовал ранее охарактеризованному белку LEA из высушенного A. avenae [9]. Кроме того, мы также идентифицировали множественные копии цитохрома Р450, супероксиддисмутазы, глутатионпероксидазы и глутатион-S-трансферазы, ферментов, участвующих в защите от окислительного повреждения. Стресс высыхания нематод вызывал значительную активацию транскриптов, кодирующих эти гены [52, 53].
Стресс высыхания нематод вызывал значительную активацию транскриптов, кодирующих эти гены [52, 53].
Функциональная классификация, основанная на генной онтологии
Генная онтология (ГО) широко используется для предсказания функции и классификации генов [54].BLAST2GO [55, 56], универсальный веб-инструмент аннотирования, использовался для назначения терминов GO для кластерных последовательностей A. avenae , извлекая их из каждого попадания BLAST против Swiss-Prot, полученного путем сопоставления с существующими ассоциациями аннотаций. 1222 последовательности из 2700 не дали никаких результатов BLAST в пределах установленного порога E-значения (<1e-5). Картирование терминов GO и аннотаций было невозможно для 173 и 205 последовательностей соответственно. Остальные 1100 (40,7%) последовательностей были успешно аннотированы и сопоставлены с одним или несколькими из трех принципов организации GO: биологический процесс, молекулярная функция и клеточный компонент.Совпадения, полученные в результате этого анализа, суммированы на фиг. 4A-4C.
4A-4C.
Резюме аннотации онтологии генов, присвоенной BLAST2GO . (A) Наиболее представленные термины GO (по количеству представленных последовательностей) основной категории «биологический процесс»; (B) Наиболее представленные термины GO основной категории «молекулярная функция» (C) Наиболее представленные термины GO основной категории «клеточный компонент». Многоуровневые круговые диаграммы были созданы с использованием отсечений последовательности 140, 50 и 40 для биологический процесс», «молекулярная функция» и «клеточный компонент» соответственно.
1003 из кластерных последовательностей A. avenae генерировали совпадения в классе «молекулярная функция», 933 в классе «биологический процесс» и 924 в классе «клеточный компонент». В классе «биологический процесс» наиболее представлены категории «регуляция биологического процесса (GO:0050789)», «биосинтетический процесс (GO:0009058)» и «транспорт (GO:0006810)», за которыми следуют категории «реакция на стимул ( GO:0050896)», на основе аннотации, присвоенной BLAST2GO (рис. 4А).В классе «молекулярная функция» наиболее представлен термин «связывание с белком (GO:0005515)», за которым следуют «связывание нуклеотидов (GO:0000166)» и «трансферазная активность (GO:0016740)» (фиг. 4B). Многие кластерные последовательности, кодирующие рибосомные белки, а также высоко экспрессированные гены, кодирующие структурные молекулы (такие как актин) и регуляторные молекулы (такие как факторы транскрипции), относятся к термину «связывание с белком». Поскольку эти кластеры в изобилии присутствуют в наборе данных, это может привести к чрезмерному представлению термина «связывание с белком».В классе «клеточный компонент» наиболее широко представлена «митохондрия (GO: 0005739)» (рис. 4C). Полный список карт GO, присвоенных кластерным последовательностям A. avenae , приведен в дополнительном файле 3.
4А).В классе «молекулярная функция» наиболее представлен термин «связывание с белком (GO:0005515)», за которым следуют «связывание нуклеотидов (GO:0000166)» и «трансферазная активность (GO:0016740)» (фиг. 4B). Многие кластерные последовательности, кодирующие рибосомные белки, а также высоко экспрессированные гены, кодирующие структурные молекулы (такие как актин) и регуляторные молекулы (такие как факторы транскрипции), относятся к термину «связывание с белком». Поскольку эти кластеры в изобилии присутствуют в наборе данных, это может привести к чрезмерному представлению термина «связывание с белком».В классе «клеточный компонент» наиболее широко представлена «митохондрия (GO: 0005739)» (рис. 4C). Полный список карт GO, присвоенных кластерным последовательностям A. avenae , приведен в дополнительном файле 3.
Идентификация транскриптов, кодирующих ферменты, разрушающие клеточную стенку кодирующие ферменты, разрушающие клеточные стенки растений и грибов (табл. 4). Идентифицированные ферменты, разрушающие клеточную стенку растений, включали целлюлазу, пектатлиазу, полигалактуроназу и экспансин, тогда как транскрипты, кодирующие ферменты, разрушающие клеточную стенку грибов, включали β-1,3-эндоглюканазу и хитиназу.
 Таблица 4 Транскрипты A. avenae , сходные с ферментами, разрушающими клеточную стенку растений или грибов
Таблица 4 Транскрипты A. avenae , сходные с ферментами, разрушающими клеточную стенку растений или грибов Восемь генов целлюлаз присутствовали в 8 различных кластерах A. avenae , и во всех случаях гомологи были обнаружены у других нематод, паразитирующих на растениях. Один ген целлюлазы (AAC01152) был идентифицирован как совокупность шести отдельных EST. Два кластера (AAC00199 и AAC00801) содержали по 2 EST каждый, а остальные кластеры целлюлазы присутствовали в виде одного элемента. Были идентифицированы два типа ферментов, разлагающих пектин: пектатлиаза и полигалактуроназа (табл. 4).В то время как все транскрипты, кодирующие гены пектатлиазы, были идентифицированы как одиночные, кластеры полигалактуроназ содержали либо одиночные, либо два индивидуальных EST. Особенности последовательностей целлюлазы и пектатлиазы более подробно обсуждаются ниже.
Помимо ферментов, разрушающих клеточную стенку растений, мы идентифицировали гены, кодирующие экспансиноподобные белки, в наборе данных A. avenae . Экспансины и экспансиноподобные белки недавно были описаны у нескольких нематод, паразитирующих на растениях [19, 31–33], и считается, что эти белки разрушают нековалентные связи в клеточной стенке растений, усиливая активность других ферментов, таких как целлюлазы.Все четыре экспансиноподобных транскрипта были идентифицированы как наиболее соответствующие генам экспансина из B. xylophilus [19].
avenae . Экспансины и экспансиноподобные белки недавно были описаны у нескольких нематод, паразитирующих на растениях [19, 31–33], и считается, что эти белки разрушают нековалентные связи в клеточной стенке растений, усиливая активность других ферментов, таких как целлюлазы.Все четыре экспансиноподобных транскрипта были идентифицированы как наиболее соответствующие генам экспансина из B. xylophilus [19].
Были идентифицированы два разных типа генов, β-1,3-эндоглюканаза и хитиназа, которые могут кодировать ферменты, важные для деградации клеточной стенки грибов. Ген, кодирующий β-1,3-эндоглюканазу, был клонирован и охарактеризован из B. xylophilus [17], и считается, что он способствует питанию этой нематоды грибами. Хитиназа, фермент, ответственный за разрушение β-1,4-гликозидных связей в хитине, была обнаружена у широкого круга нематод.Поскольку известно, что хитин присутствует в яичной скорлупе [57] и микрофилярной оболочке [58] нематод, было высказано предположение, что хитиназы играют роль в процессах ремоделирования во время линьки филярий и в вылуплении личинок из яичной скорлупы. 59, 60]. Однако наличие больших семейств хитиназ у свободноживущей нематоды C. elegans предполагает, что эти ферменты могут выполнять и другие функции [61]. Было обнаружено, что у нематоды, паразитирующей на растениях, Heterodera glycines , хитиназа экспрессируется в клетках субвентральной пищеводной железы паразитических стадий этой нематоды, что указывает на роль в паразитизме, но не в вылуплении [62].Паразитическая фитофага B. xylophilus также содержит хитиназу [15]. Поскольку β-1,3-глюкан и хитин являются двумя основными структурными полисахаридами клеточной стенки грибов, возможно, что питающиеся грибами нематоды, такие как B. xylophilus и A. avenae , выделяют эти ферменты для метаболизма или смягчают клеточную стенку грибка в процессе питания.
59, 60]. Однако наличие больших семейств хитиназ у свободноживущей нематоды C. elegans предполагает, что эти ферменты могут выполнять и другие функции [61]. Было обнаружено, что у нематоды, паразитирующей на растениях, Heterodera glycines , хитиназа экспрессируется в клетках субвентральной пищеводной железы паразитических стадий этой нематоды, что указывает на роль в паразитизме, но не в вылуплении [62].Паразитическая фитофага B. xylophilus также содержит хитиназу [15]. Поскольку β-1,3-глюкан и хитин являются двумя основными структурными полисахаридами клеточной стенки грибов, возможно, что питающиеся грибами нематоды, такие как B. xylophilus и A. avenae , выделяют эти ферменты для метаболизма или смягчают клеточную стенку грибка в процессе питания.
Характеристика генов, кодирующих целлюлазы и пектатлиазы; анализ последовательностей, филогенетика и анализ экспрессии
Целлюлаза и пектатлиаза идентифицированы и охарактеризованы у широкого круга нематод, паразитирующих на растениях [16, 18, 20–29]. Наличие генов, кодирующих ферменты, разрушающие клеточную стенку, у A. avenae , открывает новые возможности для дальнейших молекулярных исследований, направленных на понимание их функциональной роли у этой нематоды и изучение происхождения и эволюции этих генов у нематод. Поэтому мы клонировали полноразмерные кДНК и геномные последовательности двух предполагаемых целлюлаз (названных Aa-eng-1 и Aa-eng-2 ) и двух предполагаемых пектатлиаз (названных Aa-pel-1 и Aa- pel-2 ) и сравнили эти последовательности с последовательностями других нематод, паразитирующих на растениях.
Наличие генов, кодирующих ферменты, разрушающие клеточную стенку, у A. avenae , открывает новые возможности для дальнейших молекулярных исследований, направленных на понимание их функциональной роли у этой нематоды и изучение происхождения и эволюции этих генов у нематод. Поэтому мы клонировали полноразмерные кДНК и геномные последовательности двух предполагаемых целлюлаз (названных Aa-eng-1 и Aa-eng-2 ) и двух предполагаемых пектатлиаз (названных Aa-pel-1 и Aa- pel-2 ) и сравнили эти последовательности с последовательностями других нематод, паразитирующих на растениях.
Полноразмерные последовательности целлюлаз были идентифицированы из двух плазмидных клонов, чьи последовательности EST являются частью кластера AAC001152 (таблица 4). Хотя шесть отдельных EST образуют контиг для представления этого кластера, был выбран один EST, так как он показал немного отличающуюся нуклеотидную последовательность от других пяти EST. Два разных плазмидных клона, содержащих полноразмерные последовательности кДНК целлюлаз, впоследствии отбирали и секвенировали с использованием конкретных праймеров, перечисленных в таблице 5.
кДНК Aa-eng-1 имела длину 1104 п.н. (исключая полиА-хвост) и включала открытую рамку считывания (ORF) длиной 981 п.н. который может кодировать белок из 327 аминокислот со стартовым кодоном ATG в положении 35 и стоп-кодоном TGA в положении 1016 (рис. 5). Полная кДНК Aa-eng-2 имела длину 1107 п.н. и также содержала потенциальную ORF из 327 аминокислот со стартовым кодоном ATG в положении 35 и стоп-кодоном TGA в положении 1016.Сигнальный пептид из 19 аминокислот предсказан SignalP [63] на N-конце выведенных полипептидов AA-ENG-1 и AA-ENG-2. Предсказанная молекулярная масса предполагаемых зрелых белков составляла 34,130 кДа и 34,059 кДа соответственно, а теоретическое значение pI составляло 6,2 для обоих белков. Белки AA-ENG-1 и AA-ENG-2 содержали каталитический домен, гомологичный β-1,4-эндоглюканазам GHF5, как предсказано PRODOM [64]. Выведенные аминокислотные последовательности показали наибольшее сходство с эндоглюканазой GHF5 из мигрирующей растительной паразитической нематоды Radopholus similis (GenBank Accession No.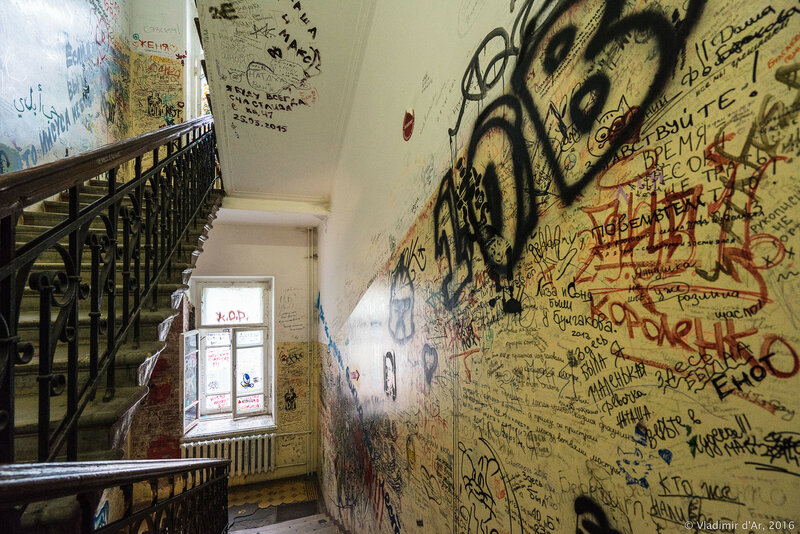 [ABV54446]). Наибольшее нематодное сходство как AA-ENG-1, так и AA-ENG-2 было с β-1,4-эндоглюканазой из Cytophaga hutchinsoni (целлюлолитическая скользящая бактерия; инвентарный номер GenBank [YP_678708]). Аминокислотные последовательности AA-ENG-1 и AA-ENG-2 совпадают на 99%.
[ABV54446]). Наибольшее нематодное сходство как AA-ENG-1, так и AA-ENG-2 было с β-1,4-эндоглюканазой из Cytophaga hutchinsoni (целлюлолитическая скользящая бактерия; инвентарный номер GenBank [YP_678708]). Аминокислотные последовательности AA-ENG-1 и AA-ENG-2 совпадают на 99%.
Aa-eng-1 Последовательность кДНК и предсказанные аминокислотные последовательности . Предсказанный сигнальный пептид для секреции и сигнальная последовательность полиаденилирования подчеркнуты.Предсказанные положения пяти идентифицированных последовательностей интронов обозначены затемненными треугольниками. Праймеры, используемые для получения полноразмерной последовательности кДНК и геномной амплификации, указаны стрелками. Аминокислоты в прямоугольниках представляют предполагаемые остатки активного сайта.
Геномные клоны Aa-eng-1 и Aa-eng-2 были получены с помощью ПЦР-амплификации с использованием геноспецифических праймеров (таблица 5) и геномной ДНК в качестве матрицы. Продукты геномной ДНК Aa-eng-1 и Aa-eng-2 имели длину 1518 п.н. и 1522 п.н. соответственно от ATG до стоп-кодона.Положение границ экзона/интрона геномных последовательностей определяли путем сопоставления геномных последовательностей с соответствующими последовательностями кДНК. Все интроны окаймлены каноническими последовательностями цис-сплайсинга [65]. Пять интронов были идентифицированы у Aa-eng-1 (рис. 5), из которых четыре интрона были небольшими (от 40 до 85 п.н.), что обычно встречается у нематод [66]. Только первый интрон был крупнее (319 п.н.). Пять интронов были также идентифицированы в Aa-eng-2 . Первый интрон имел длину 337 п.н., а длина остальных четырех интронов колебалась от 40 до 72 п.н.Положения интронов генов Aa-eng-1 и Aa-eng-2 идентичны друг другу.
Продукты геномной ДНК Aa-eng-1 и Aa-eng-2 имели длину 1518 п.н. и 1522 п.н. соответственно от ATG до стоп-кодона.Положение границ экзона/интрона геномных последовательностей определяли путем сопоставления геномных последовательностей с соответствующими последовательностями кДНК. Все интроны окаймлены каноническими последовательностями цис-сплайсинга [65]. Пять интронов были идентифицированы у Aa-eng-1 (рис. 5), из которых четыре интрона были небольшими (от 40 до 85 п.н.), что обычно встречается у нематод [66]. Только первый интрон был крупнее (319 п.н.). Пять интронов были также идентифицированы в Aa-eng-2 . Первый интрон имел длину 337 п.н., а длина остальных четырех интронов колебалась от 40 до 72 п.н.Положения интронов генов Aa-eng-1 и Aa-eng-2 идентичны друг другу.
Выравнивание последовательностей двух эндоглюканаз из A. avenae с эндоглюканазами GHF5 из нематод и других организмов показало, что как AA-ENG-1, так и AA-ENG-2 обладают консенсусным паттерном эндоглюканаз GHF5 в их первичных аминокислотных последовательностях в какие два остатка глутаминовой кислоты являются предполагаемым донором протона и муклеофилом/основанием каталитического центра (рис. 5), что справедливо и для всех ранее описанных эндоглюканаз GHF5 нематод. В дополнение к каталитическому домену некоторые из этих белков содержат целлюлозосвязывающий домен (CBD), соединенный с каталитическим доменом через линкерный пептид. Гены эндоглюканазы GHF5, выделенные из нематод, паразитирующих на растениях, также имеют различную структуру: все имеют сигнальный пептид и каталитический домен, некоторые имеют дополнительный линкер и CBD, а другие имеют только линкер, но не CBD [25, 67]. Однако ни пептидные линкеры, ни домены CBD не присутствовали в двух эндоглюканазах GHF5, выделенных из A.авеню . Также было показано, что экспансины из цистообразующих нематод содержат CBD, но такие домены не были предсказаны в других последовательностях в наборе данных EST, включая предполагаемые экспансины.
5), что справедливо и для всех ранее описанных эндоглюканаз GHF5 нематод. В дополнение к каталитическому домену некоторые из этих белков содержат целлюлозосвязывающий домен (CBD), соединенный с каталитическим доменом через линкерный пептид. Гены эндоглюканазы GHF5, выделенные из нематод, паразитирующих на растениях, также имеют различную структуру: все имеют сигнальный пептид и каталитический домен, некоторые имеют дополнительный линкер и CBD, а другие имеют только линкер, но не CBD [25, 67]. Однако ни пептидные линкеры, ни домены CBD не присутствовали в двух эндоглюканазах GHF5, выделенных из A.авеню . Также было показано, что экспансины из цистообразующих нематод содержат CBD, но такие домены не были предсказаны в других последовательностях в наборе данных EST, включая предполагаемые экспансины.
Филогенетическое дерево было создано путем выравнивания белковых последовательностей β-1,4-эндоглюканазы из AA-ENG-1, AA-ENG-2, цисто- и галловых нематод, мигрирующих нематод, паразитирующих на растениях R. similis, Pratylenchus penetrans, Pratylenchus coffeae и Ditylenchus africanus и целлюлазы GHF5 из жуков-фитофагов, бактерий и простейших (рис.6). AA-ENG-1 и AA-ENG-2 объединены в более крупную группу белковых последовательностей, включающую все целлюлазы GHF5 нематод, что указывает на то, что целлюлазы A. avenae тесно связаны с целлюлазами Tylenchida. Этот анализ подтверждает идею о том, что все целлюлазы нематод GHF5 произошли от последовательности GHF5, приобретенной общим предком этой группы.
similis, Pratylenchus penetrans, Pratylenchus coffeae и Ditylenchus africanus и целлюлазы GHF5 из жуков-фитофагов, бактерий и простейших (рис.6). AA-ENG-1 и AA-ENG-2 объединены в более крупную группу белковых последовательностей, включающую все целлюлазы GHF5 нематод, что указывает на то, что целлюлазы A. avenae тесно связаны с целлюлазами Tylenchida. Этот анализ подтверждает идею о том, что все целлюлазы нематод GHF5 произошли от последовательности GHF5, приобретенной общим предком этой группы.
Неукорененное филогенетическое дерево каталитических доменов GHF5 на основе белковых последовательностей с использованием метода максимального правдоподобия .Белки GHF5 из A. avenae (AA-ENG-1 и AA-ENG-2) выделены жирным шрифтом. Номера доступа GenBank белков GHF5 из Meloidogyne incognita (MI-ENG-1, MI-ENG-2, MI-ENG-3 и MI-ENG-4), Pratylenchus penetrans (PP-ENG-1 и PP- ENG-2), Pratylenchus coffeae (PC-ENG-1), Radopholus similis (RS-ENG-1A, RS-ENG-1B, RS-ENG-2 и RS-ENG-3), Globodera rostochiensis (GR-ENG-1, GR-ENG-2 и GR-ENG-4), Heterodera глицин (HG-ENG-2, HG-ENG-4, HG-ENG-5 и HG-ENG-6) , Ditylenchus africanus (DA-ENG-1), в скобках указаны жуки, бактерии и протисты. Начальные значения рассчитываются из 1000 повторов. Шкала шкалы представляет 10 замен на 100 позиций аминокислот.
Начальные значения рассчитываются из 1000 повторов. Шкала шкалы представляет 10 замен на 100 позиций аминокислот.
β-1,4-эндоглюканазы представляют собой крупнейшее семейство ферментов, разрушающих клеточную стенку, которые на сегодняшний день идентифицированы у паразитических нематод. За последнее десятилетие было идентифицировано и широко изучено большое количество эндоглюканаз GHF5 у паразитических растений Tylenchida, включая цистовые и галловые нематоды [20–25]. Гены, кодирующие β-1,4-эндоглюканазы, также обнаружены у Bursaphelenchus spp, но эти ферменты наиболее сходны с целлюлазами GHF45 грибов [16].Присутствие целлюлаз GHF5 в составе A. avenae (в отличие от целлюлаз GHF45) подтверждает предположение, что эта нематода более тесно связана с Tylenchida, чем с Bursaphelenchus и его родственниками.
Хотя присутствие целлюлаз GHF5 в A. avenae можно легко объяснить, учитывая приведенные выше филогенетические аргументы, присутствие β-1,3-эндоглюканазы более удивительно. Эти ферменты метаболизируют клеточную стенку грибов и ранее были описаны у Bursaphelenchus spp.Такие гены обычно не присутствуют у животных, и было высказано предположение, что генов Bursaphelenchus были приобретены путем горизонтального переноса генов от бактерий [17].
Эти ферменты метаболизируют клеточную стенку грибов и ранее были описаны у Bursaphelenchus spp.Такие гены обычно не присутствуют у животных, и было высказано предположение, что генов Bursaphelenchus были приобретены путем горизонтального переноса генов от бактерий [17].
Такие гены отсутствуют у галловых нематод (для которых доступны две последовательности генома) или других тиленхид. Вполне возможно, что общий предок Aphelenchoidea и Tylenchida, питающийся грибами, обладал этим геном, но впоследствии его утратили более «продвинутые» растительные паразиты. Для решения этой проблемы требуется дальнейшее секвенирование в обеих группах нематод.
Все транскрипты, потенциально кодирующие пектатлиазы, были идентифицированы как синглетоны (табл. 4). Две полноразмерные последовательности кДНК пектатлиаз, обозначенные как Aa-pel-1 и Aa-pel-2 , были идентифицированы из плазмидных клонов, соответствующих идентификаторам кластера AAC01649 и AAC03048 соответственно. Полная кДНК Aa-pel-1 имела длину 821 п.н. и содержала ORF из 247 аминокислот с предполагаемым стартовым кодоном ATG в положении 30 и стоп-кодоном TAA в положении 771.Сигнальный пептид из 18 аминокислот предсказан SignalP [63] на N-конце предполагаемой аминокислотной последовательности AA-PEL-1. Зрелый белок имеет расчетную молекулярную массу 24,250 кДа и теоретическое pI 8,93.
Полная кДНК Aa-pel-1 имела длину 821 п.н. и содержала ORF из 247 аминокислот с предполагаемым стартовым кодоном ATG в положении 30 и стоп-кодоном TAA в положении 771.Сигнальный пептид из 18 аминокислот предсказан SignalP [63] на N-конце предполагаемой аминокислотной последовательности AA-PEL-1. Зрелый белок имеет расчетную молекулярную массу 24,250 кДа и теоретическое pI 8,93.
Полноразмерная кДНК Aa-pel-2 имела длину 838 п.н. и содержала ORF длиной 249 п.н. со стартовым кодоном ATG в положении 31 и стоп-кодоном TAA в положении 778. N-концевая сигнальная последовательность из 19 аминокислот, предсказанных SignalP [63]. Молекулярная масса и теоретическое значение pI предполагаемого белка AA-PEL-2 составляли 24.329 кДа и 9,12 соответственно. AA-PEL-1 на 61% идентичен AA-PEL-2.
Для получения геномных последовательностей все кодирующие области гена Aa-pel-1 и Aa-pel-2 амплифицировали из гДНК A. avenae с использованием генспецифических праймеров (таблица 5). Анализ этих последовательностей показал, что гены Aa-pel-1 и Aa-pel-2 имели длину 1860 и 1221 п.н. соответственно от ATG до стоп-кодона. Два интрона (468 п.н. и 651 п.н.) были идентифицированы в Aa-pel-1 , тогда как Aa-pel-2 содержали только один интрон (690 п.н.) (рис.7). Все интроны окаймлены каноническими последовательностями цис-сплайсинга [65]. Положение второго интрона Aa-pel-1 было идентично интрону Aa-pel-2 . Рис. 7 и грибок . BX-PEL-1 [BAE48369], BX-PEL-2 [BAE48370] из Bursaphelenchus xylophilus ; BM-PEL-1 [BAE48373] и BM-PEL-2 [BAE48375] из Bursaphelenchus mucronatus ; MI-PEL-1 [AAQ09004] и MI-PEL-2 [AAQ97032] из Meloidogyne incognita ; MJ-PEL-1 [AAL66022] из Meloidogyne javanica ; GR-PEL-1 [AAF80747] из Globodera rostochiensis ; SC-PEL-1 [NP625403] из Streptomyces coelicolor и FS-PEL-B [AAA8738] из Fusarium solani .Идентичные остатки выделены черным цветом, а функционально консервативные — серым.
Анализ этих последовательностей показал, что гены Aa-pel-1 и Aa-pel-2 имели длину 1860 и 1221 п.н. соответственно от ATG до стоп-кодона. Два интрона (468 п.н. и 651 п.н.) были идентифицированы в Aa-pel-1 , тогда как Aa-pel-2 содержали только один интрон (690 п.н.) (рис.7). Все интроны окаймлены каноническими последовательностями цис-сплайсинга [65]. Положение второго интрона Aa-pel-1 было идентично интрону Aa-pel-2 . Рис. 7 и грибок . BX-PEL-1 [BAE48369], BX-PEL-2 [BAE48370] из Bursaphelenchus xylophilus ; BM-PEL-1 [BAE48373] и BM-PEL-2 [BAE48375] из Bursaphelenchus mucronatus ; MI-PEL-1 [AAQ09004] и MI-PEL-2 [AAQ97032] из Meloidogyne incognita ; MJ-PEL-1 [AAL66022] из Meloidogyne javanica ; GR-PEL-1 [AAF80747] из Globodera rostochiensis ; SC-PEL-1 [NP625403] из Streptomyces coelicolor и FS-PEL-B [AAA8738] из Fusarium solani .Идентичные остатки выделены черным цветом, а функционально консервативные — серым. Черные столбцы (от I до IV) указывают на консервативные области, характерные для пектатлиаз PL3. Очень высококонсервативные заряженные остатки обозначены под выравниванием числовым символом (#), а звездочка (*) указывает на консервативные цистеиновые остатки. Положения интрона в AA-PEL-1 и AA-PEL-2 обозначены A1 и A2 соответственно, а в BX-PEL-1/2 и BM-PEL-1/2 — B . , а те в МИ-РУС-1 по Ми1 .Треугольники и ромбы представляют интроны фазы 0 и 1 соответственно.
Черные столбцы (от I до IV) указывают на консервативные области, характерные для пектатлиаз PL3. Очень высококонсервативные заряженные остатки обозначены под выравниванием числовым символом (#), а звездочка (*) указывает на консервативные цистеиновые остатки. Положения интрона в AA-PEL-1 и AA-PEL-2 обозначены A1 и A2 соответственно, а в BX-PEL-1/2 и BM-PEL-1/2 — B . , а те в МИ-РУС-1 по Ми1 .Треугольники и ромбы представляют интроны фазы 0 и 1 соответственно.
Положение интрона в генах A. avenae ( Aa-pel-1 и Aa-pel-2 ) сравнивали с другими генами пектатлиазы нематод. Гены пектатлиазы из видов Bursaphelenchus ( Bx-pel-1/2 и Bm-pel-1/2 ) имеют по одному интрону в своей кодирующей области в одном и том же положении [18]. Это положение также идентично общему положению интрона A.avenae генов (рис. 7). Более того, один из трех интронов Mi-pel-1 из M. incognita находится в том же положении, что и в генах A. avenae и Bursaphelenchus . Gr-pel-1 из G. rostochiensis имеет шесть интронов, а Mi-pel-2 из M. incognita имеет два интрона. Gr-pel-1 и Mi-pel-2 имеют два общих положения интрона, но ни один из интронов этих генов не имеет того же положения, что и в A.avenae генов (не показаны).
avenae и Bursaphelenchus . Gr-pel-1 из G. rostochiensis имеет шесть интронов, а Mi-pel-2 из M. incognita имеет два интрона. Gr-pel-1 и Mi-pel-2 имеют два общих положения интрона, но ни один из интронов этих генов не имеет того же положения, что и в A.avenae генов (не показаны).
Поиск гомологии белка с использованием выведенных аминокислотных последовательностей AA-PEL-1 и AA-PEL-2 с использованием BLASTP показал высокое сходство с пектатлиазами, принадлежащими к семейству полисахаридлиаз 3 (PL3) из растительных паразитических нематод, бактерий и грибы. Множественное выравнивание последовательностей AA-PEL-1 и AA-PEL-2 с лучшим совпадением подтвердило, что обе последовательности A. avenae содержат четыре высококонсервативных участка, характерных для пектатлиаз PL3 в бактериях и грибах, а также 8-10 цистеина. остатков и четыре заряженных остатка (рис.7), которые потенциально участвуют в катализе [26, 68]. AA-PEL-1 и AA-PEL-2 были наиболее сходны с пектатлиазами (BX-PEL1/2 и BM-PEL1/2) сосновых нематод B. xylophilus и B. mucronatus (от 52 до 59). % идентичности для AA-PEL-1 и от 53 до 63% идентичности для AA-PEL-2) [18]. Последовательности A. avenae имеют от 23 до 33% идентичности с пектатлиазами (MI-PEL-1, MI-PEL-2, MJ-PEL-1 и GR-PEL-1) из цист и корневых узлов растений, паразитирующих нематодами spp. [28, 29], и от 39 до 45% идентичности с последовательностями двух микробов (рис.7).
AA-PEL-1 и AA-PEL-2 были наиболее сходны с пектатлиазами (BX-PEL1/2 и BM-PEL1/2) сосновых нематод B. xylophilus и B. mucronatus (от 52 до 59). % идентичности для AA-PEL-1 и от 53 до 63% идентичности для AA-PEL-2) [18]. Последовательности A. avenae имеют от 23 до 33% идентичности с пектатлиазами (MI-PEL-1, MI-PEL-2, MJ-PEL-1 и GR-PEL-1) из цист и корневых узлов растений, паразитирующих нематодами spp. [28, 29], и от 39 до 45% идентичности с последовательностями двух микробов (рис.7).
Филогенетическое дерево было создано путем сопоставления последовательностей пектатлиазы A. avenae с выбранными белками, принадлежащими к PL3, из бактерий, грибов и нематод с использованием метода максимального правдоподобия (рис. 8). Оба Aa-pel-1 и Aa-pel-2 были сгруппированы с генами Bursaphelenchus . Другие последовательности нематод не были монофилетическими, а были сгруппированы в отдельные клады. Рис. 8Белки PL3 из A. avenae (AA-PEL-1 и AA-PEL-2) выделены жирным шрифтом. Номера доступа GenBank белков PL3 из Bursaphelenchus mucronatus (BM-PEL-1 и BM-PEL-2), Bursaphelenchus xylophilus (BX-PEL-1 и BX-PEL-2), Meloidogyne incognita (MI- PEL-1, MI-PEL-2 и MI-PEL-3), Meloidogyne javanica (MJ-PEL), Globodera rostochiensis (GR-PEL-1 и GR-PEL-2), Heterodera schachtii ( HS-PEL-1 и HS-PEL-2), в скобках указаны бактерии и грибы.Начальные значения рассчитываются из 1000 повторов. Шкала шкалы представляет 20 замен на 100 позиций аминокислот.
Номера доступа GenBank белков PL3 из Bursaphelenchus mucronatus (BM-PEL-1 и BM-PEL-2), Bursaphelenchus xylophilus (BX-PEL-1 и BX-PEL-2), Meloidogyne incognita (MI- PEL-1, MI-PEL-2 и MI-PEL-3), Meloidogyne javanica (MJ-PEL), Globodera rostochiensis (GR-PEL-1 и GR-PEL-2), Heterodera schachtii ( HS-PEL-1 и HS-PEL-2), в скобках указаны бактерии и грибы.Начальные значения рассчитываются из 1000 повторов. Шкала шкалы представляет 20 замен на 100 позиций аминокислот.
Гены пектатлиазы из A. avenae более похожи на гены Bursaphelenchus по сравнению с генами цисто- и галловых нематод (рис. 7 и 8). Идентичное положение общего интрона генов A. avenae ( Aa-pel-1 и Aa-pel-2 ) и интронов в составе генов пектатлиазы Bursaphelenchus и M.incognita (рис. 7) позволяет предположить, что гены пектатлиазы широкого круга нематод, паразитирующих на растениях, имеют одинаковое происхождение. Чтобы определить, какие клетки A. avenae экспрессируют эндоглюканазы GHF5 и пектатлиазы, проводили гибридизацию in situ мРНК (фиг. 9). Меченные дигоксигенином антисмысловые зонды, полученные из Aa-eng-1 и Aa-pel-1 , специфически гибридизировались с транскриптами в клетках пищеводной железы A. avenae (фиг.9А и 9С). Окрашивание наблюдалось у молодых и взрослых нематод, а не ограничивалось определенными стадиями жизни. Гибридизации с контрольными (смысловыми) кДНК-зондами Aa-eng-1 или Aa-pel-1 не наблюдалось (фиг. 9B и 9D).
Чтобы определить, какие клетки A. avenae экспрессируют эндоглюканазы GHF5 и пектатлиазы, проводили гибридизацию in situ мРНК (фиг. 9). Меченные дигоксигенином антисмысловые зонды, полученные из Aa-eng-1 и Aa-pel-1 , специфически гибридизировались с транскриптами в клетках пищеводной железы A. avenae (фиг.9А и 9С). Окрашивание наблюдалось у молодых и взрослых нематод, а не ограничивалось определенными стадиями жизни. Гибридизации с контрольными (смысловыми) кДНК-зондами Aa-eng-1 или Aa-pel-1 не наблюдалось (фиг. 9B и 9D).
Локализация AA-ENG-1
и 2 и AA-PEL-1 AA-PEL-1 Стенограммы в клетки пищевода в ячейках эзофагеации A. Avenae на in situ гибридизация .Срезы нематод гибридизовали с антисмысловыми (А) и смысловыми (В) зондами кДНК, меченными дигоксигенином Aa-eng-1 . Гибридизацию также проводили с мечеными дигоксигенином антисмысловыми (С) и смысловыми (D) зондами кДНК Aa-pel-1 . G, пищеводные железы; S, стилет; М, метакорпус. Полоса указывает длину 20 мкм.
Гибридизацию также проводили с мечеными дигоксигенином антисмысловыми (С) и смысловыми (D) зондами кДНК Aa-pel-1 . G, пищеводные железы; S, стилет; М, метакорпус. Полоса указывает длину 20 мкм. В рамках сложного процесса паразитизма широкий спектр нематод, паразитирующих на растениях, использует эндогенные β-1,4-эндоглюканазы и пектатлиазы для деградации двух распространенных компонентов клеточной стенки растений и, таким образом, облегчения их миграции через ткани хозяина.Присутствие сигнальных пептидов в выведенных аминокислотных последовательностях эндоглюканаз и пектатлиазы из A. avenae в сочетании с их экспрессией в пищеводных железах предполагает, что оба фермента играют аналогичную роль в A. avenae . Наличие таких генов у A. avenae предполагает, что эта нематода может проникать и мигрировать через ткани растений, а также питаться содержимым клеток растений. Это подтверждается наблюдением, что A. avenae , как известно, питается тканями растений в культуре [13, 14]. Таким образом, A. avenae может иметь разнообразную диету, включающую грибы и растительные ткани. Это, в сочетании с положением этой нематоды как основного члена клады, которая включает широкий спектр нематод, паразитирующих на растениях, обеспечивает дополнительную поддержку идеи о том, что паразитизм растений развился в результате питания грибами, и предполагает, что A. avenae , может быть очень примитивным питателем растений.
Таким образом, A. avenae может иметь разнообразную диету, включающую грибы и растительные ткани. Это, в сочетании с положением этой нематоды как основного члена клады, которая включает широкий спектр нематод, паразитирующих на растениях, обеспечивает дополнительную поддержку идеи о том, что паразитизм растений развился в результате питания грибами, и предполагает, что A. avenae , может быть очень примитивным питателем растений.
10 хитрых способов повесить подарочные бирки на подарки для вечеринок
Зарегистрируйтесь здесь, чтобы стать нашим оптовым продавцом подарочных бирок и наклеек.Создайте для себя задачу по изготовлению предметов, используя подарочные жетоны. Не вешайте их просто на веревке. Есть много способов прикрепить подарочные бирки, чтобы превратить ваши подарки в предмет для разговора. Подарочные ярлыки также предлагают простой способ украсить ваши свадебные сувениры.
Использование подарочных ярлыков — это простой и отличный способ украсить ваши подарки. Кроме того, вы можете отправить короткое сердечное послание на подарочной бирке вместо объемной поздравительной открытки. Самый простой способ прикрепить подарочную бирку к подарочной коробке — повесить ее на веревке.Но мы хотим показать вам более творческие и веселые способы сделать это.
Кроме того, вы можете отправить короткое сердечное послание на подарочной бирке вместо объемной поздравительной открытки. Самый простой способ прикрепить подарочную бирку к подарочной коробке — повесить ее на веревке.Но мы хотим показать вам более творческие и веселые способы сделать это.
Мы также хотим продемонстрировать, как небольшие ремесленные навыки могут улучшить подарочную бирку. Просто посмотрите видеоуроки, которые мы создали ниже.
1. Мини-баночки и палочки для мороженого Nutella
Источник: giftelements.comМини-баночки Nutella или мини-баночки для варенья — отличный подарок на день рождения или свадьбу. Nutella — такой популярный бренд, который сам по себе привлекает внимание.Вы буквально чувствуете вкус шоколада во рту при виде баночки с Nutella. Поэтому следите за тем, чтобы оформление подарка не мешало новизне бренда.
Подарочная бирка крепится на палочке от мороженого и приклеивается к задней части банки. Таким образом, впечатления от подарочной бирки и бренда продукта будут дополнять друг друга, а не конкурировать. Добавьте больше стиля, украсив ленточками и бантами, чтобы завершить образ.
Добавьте больше стиля, украсив ленточками и бантами, чтобы завершить образ.
2. Наклейте подарок на подарочную бирку
Ссылка на пинтерест: https://пин.it/LXPB5CYHQJPX5NPинтересная ссылка: https://pin.it/TC5XEP6ZJS2FT7Эта идея представляет собой интересный подход к смене ролей. В наши дни смена ролей в браке совершенно нормальна. Но что, если подарок прикреплен к подарочной бирке, а не наоборот? Теперь это инновационно. Это такой умный способ добавить размер к крошечным подаркам, таким как эти одиночные кусочки шоколада или даже маленькие сувениры.
Убедитесь, что подарочные бирки изготовлены из более прочного картона, чтобы они не провисали и не сгибались.Если подарок преподносят в качестве сувенира для вечеринки, привяжите подарочные бирки к стеклянной посуде или столовым приборам, чтобы их не разбрасывали.
3. Многослойные подарочные бирки
Ссылка на Pinterest: https://pin.it/OHYUXJ3JZODVKTPСсылка на Pinterest: https://pin.it/4ROXWDX5X4CEOPНаслоение различных материалов на подарочную бирку может сделать ваш подарок ослепительным. Вам потребуется как минимум 2 типа бумажного материала и интерес к простой работе. Но усилия того стоят. Используйте материалы с разными текстурами, мотивами и цветами для создания трехмерного эффекта.Любая конкретная подарочная бирка может создавать различные впечатления с помощью различных методов наслоения.
Например, использование полупрозрачного белого пергамина или рисовой бумаги на ярком фоне может создать утонченный и романтичный образ. Или используйте маленькую полоску на большой подарочной бирке, чтобы создать эффект подвески. Вдохните жизнь в простую подарочную бирку, соединив ее с яркой красочной картой, расположенной под ней.
4. Украшения на подарочных бирках
Ссылка на пинтерест: https://пин. ссылка it/6ndPkejPinterest: https://pin.it/5LtECPl
ссылка it/6ndPkejPinterest: https://pin.it/5LtECPlЕсли у вас нет красивой подарочной бирки, чтобы украсить свой подарок, не беспокойтесь. Возьмите чистый лист бумаги и просто напишите на нем свои чувства. Если у вас не очень хороший почерк, то просто наберите и распечатайте свое сообщение. Вырежьте бумагу любой формы и размера, чтобы она соответствовала вашему подарку. Выберите травинку или веточку с листьями снаружи и приклейте ее на подарочную бирку с помощью клея или скотча.
Было бы еще лучше, если бы вы могли использовать маленький цветок.Мы предпочитаем использовать сушеные растения в качестве украшения на бирках, чтобы они не выглядели увядшими и обесцвеченными через некоторое время. Поддельные будут работать так же чудесно, чтобы придать взгляду вечную свежесть.
5.
 Перевернутые палочки для леденцов Ссылка на Pinterest: https://pin.it/6pedbpjСсылка на Pinterest: https://pin.it/6j1lmtf
Перевернутые палочки для леденцов Ссылка на Pinterest: https://pin.it/6pedbpjСсылка на Pinterest: https://pin.it/6j1lmtfЭто еще один умный способ украсить маленькие подарки и придать им роскошный вид. Ваш бюджет на вечеринку или свадьбу не должен обходиться в кругленькую сумму.Поместите круглые леденцы на палочках вверх дном в небольшую упаковку с плоским дном. Проделайте отверстия в верхней и нижней части подарочной бирки и проденьте палочку через бирку. Результат очень привлекателен для маленького творческого подвига.
Благодаря плоскому основанию леденец остается в вертикальном положении и не перекатывается. Если вы хотите пройти лишнюю милю, приобретите круглые шоколадные трюфели профессионального изготовления. Просто скажите своему пекарю или шоколатье, чтобы они добавили палочку от леденца или длинную соломинку в каждый трюфель.
6. Повесьте белье на прищепку
Ссылка на Пинтерест: https://pin.it/6HiWU5TPСсылка на Пинтерест: https://pin. it/ykw6RWF
it/ykw6RWFДеревянные прищепки для белья прекрасно дополнят подарочные бирки. Они выглядят деревенскими и улучшают стиль загородного сада. Так что замените эти ленты для галстука деревянными крючками для белья, чтобы повесить свои подарочные бирки. Обязательно проверяйте каждый колышек, прежде чем использовать их, чтобы убедиться, что сцепление прочное. Просто поместите лист бумаги между рукоятками и похлопайте им.
Если бумага не отлетает, то эта заглушка способна выполнять свою работу.Нам нравится использовать деревянные прищепки для белья, чтобы вешать подарочные ярлыки на мини-горшечные растения. Оберните горшки мешковиной, чтобы они выглядели более по-домашнему, как будто вы сами только что убрали горшки с грядки.
7. Не вешайте их, а приклеивайте
Источник: giftelements.com Подарочные ярлыки бывают всех форм, дизайнов и цветов. Первоначально подарочные бирки использовались исключительно как средство передачи сообщения с подарком. Их можно рассматривать как мини-версии поздравительных открыток, и они очень удобны для передачи кратких чувств. Но теперь подарочные бирки все чаще используются в качестве декоративных элементов для подарков и сувениров.
Но теперь подарочные бирки все чаще используются в качестве декоративных элементов для подарков и сувениров.
Самые искусные можно использовать в качестве дизайнерских панелей, дополняющих тематику подарка. Для этого закройте отверстие для шнурка на подарочной бирке украшением, например, фигурным вырезом, стразами, лентой, листиком или цветком. Используйте ленту или клей, чтобы прикрепить подарочную бирку к передней или верхней части подарка. Поместите его под небольшим углом, чтобы получить стильный наклон.
8. Закон о веревке
Ссылка на пинтерест: https://пин.it/21WRZTcPинтересная ссылка: https://pin.it/6sa3hLq Тот факт, что отверстия для подарочных ярлыков действительно крошечные, не означает, что нам нужно использовать узкие ленты, чтобы повесить их. Здесь нам нравится, как широкие ленты используются для подвешивания подарочных ярлыков. Никакой привязки не происходит. Подарочные бирки в значительной степени заперты и не будут скользить вниз, потому что широкие ленты ведут себя как стопоры. Они, безусловно, повышают уровень сложности простых подарочных пакетов.
Здесь нам нравится, как широкие ленты используются для подвешивания подарочных ярлыков. Никакой привязки не происходит. Подарочные бирки в значительной степени заперты и не будут скользить вниз, потому что широкие ленты ведут себя как стопоры. Они, безусловно, повышают уровень сложности простых подарочных пакетов.
Гости почувствуют, что выиграли главный приз.Определенно победитель здесь и стоит попробовать, если вы думаете о том, чтобы раздать свои услуги в подарочных пакетах.
9. Скажи это сургучом
Ссылка на пинтерест: https://pin.it/1Heq3sFЧувства не нужно выражать словами. Иллюстрации или символы могут использоваться для передачи осмысленных сообщений. Вместо подарочных бирок используйте сургуч, чтобы передать эти сердечные символы. Ваши подарки будут нести царственную ауру и стиль Средневековья. Приобретите сургуч в любом магазине декоративно-прикладного искусства.
10. Играть в одевалки
Источник: somethingturquoise. com
comЭти подарочные бирки в виде нагрудника очень мило подходят для розыгрыша напитков в бутылках. Эти этикетки для бутылок выглядят так, как будто они были профессионально напечатаны. Но на самом деле их очень просто сделать своими руками. Загрузите шаблон здесь, вырежьте его скромными ножницами, и он готов к использованию. Развлекайся!
Граффити: Искусство тега
Обновлено 04 октября 2016 г. 10:09:05
Тегирование, акт написания вашего граффити-имени аэрозольной краской или маркерами, является одной из самых клеветнических, неправильно понятых и преследуемых форм самовыражения.
С самого начала своего существования в борющихся кварталах Нью-Йорка и в переулках пригородов Австралии мнения художников и некоторых представителей общественности по-прежнему расходятся во мнениях относительно художественной ценности граффити.
Инфографика: «Лысый», автор граффити, на заброшенной фабрике на западе Мельбурна. (Новости ABC: Аллисса Отред)
(Новости ABC: Аллисса Отред)Граффити находит цвет в бедных кварталах Нью-Йорка
В эпоху, когда уличное искусство и граффити-росписи никогда не были более приемлемыми для общества, маркировка имеет серьезную имиджевую проблему: она ассоциируется с бандами, считается бездумным вандализмом и бросает вызов нашим представлениям о владение недвижимостью.
Даже в культуре граффити маркировка часто затмевается фресками или смешивается с другими формами граффити-меток, такими как наброски (обычно две буквы, состоящие из контура и заливки).
А если добавить к обсуждению стрит-арт, то для непосвященных разграничение тегов будет таким же туманным, как пары из баллончика с краской.
Эти факторы затрудняют разумное обсуждение происхождения, мотивации и эстетики маркировки даже с самыми непредубежденными людьми.
Нас приучили не любить навешивание ярлыков, или это действие, которое имеет мало или вообще не имеет эстетической ценности?
Инфографика: Бирки украшают запирающийся гараж в Балаклаве, пригороде Мельбурна. (Новости ABC: Аллисса Отред.)
(Новости ABC: Аллисса Отред.)Исследователь граффити из Мельбурнского университета доктор Лахлан МакДауэлл сказал, что маркировка была краеугольным камнем культуры граффити, которая возникла в бедных кварталах Нью-Йорка в конце 1960-х годов.
«Метки — это прототип фрески», — сказал он.
«Существует прямая связь между тегом, который становился все более и более декоративным, и в конечном итоге превратился в этот полноцветный выжигатель.»
В 1971 году, когда New York Times опубликовала статью о TAKI 183 — одном из самых первых зарегистрированных граффити-художников — не имело значения, что он был бедным подростком с Манхэттена.
В то время его метка была так распространена в нью-йоркском пейзаже, она была так же узнаваема, как и любой актер, чье имя горит на Бродвее.
Несмотря на высокий уровень безработицы в городе, пометки сделали славу, восхищение и уважение доступными даже для самых бедных жителей города.
И сейчас, более 40 лет спустя, маркировка по-прежнему предлагает те же награды.
«Когда вы молоды и бессильны, граффити — это простой способ — ну, не такой уж и легкий — заслужить уважение своих сверстников только собственным упорным трудом», — сказал художник-граффитист из восточного Мельбурна по имени Пол.
Инфографика: Пол, художник с востока Мельбурна, идет по переулку Балаклавы.(Новости ABC: Аллисса Отред)Карточка для туризма против незаконных каракулей
В начале 1980-х первые проблески нью-йоркского граффити появились в Австралии благодаря документальным фильмам, таким как «Войны стилей» (1983) и книгам, таким как «Искусство метро» (1984).
И эта культура быстро укоренилась, возможно, отчасти из-за политической и протестной культуры граффити, которая уже существовала в наших городах, о чем свидетельствует книга фотографа Ренни Эллиса «Австралийские граффити» (1975).
Отношение к городскому искусству значительно изменилось за последние четыре десятилетия. Там, где когда-то подавляющее большинство граффити считалось вандализмом, взрыв уличного искусства в конце 1990-х привел к большему признанию всех форм городского искусства, включая фрески-граффити. Но тегирование остается свободным от этого изменения отношения.
Там, где когда-то подавляющее большинство граффити считалось вандализмом, взрыв уличного искусства в конце 1990-х привел к большему признанию всех форм городского искусства, включая фрески-граффити. Но тегирование остается свободным от этого изменения отношения.
(Новости ABC: Аллисса Отред.)
За несколько коротких лет уличное искусство стало настолько важной частью туристического опыта Мельбурна, что теперь кажется, что ни одна поездка не обходится без того, чтобы посетители не сделали селфи на одном из знаменитых переулков города.
Многие другие города теперь, похоже, также стремятся извлечь выгоду из популярности уличного искусства среди туристов.
Но в то время как местные органы власти поощряют одни формы городского искусства за счет других, с точки зрения закона разграничение между законным и незаконным не было столь четким.
Профессор криминологии Мельбурнского университета Элисон Янг сказала, что не имеет значения, было ли это уличное искусство, граффити или маркировка, преступление считается преступлением, если работа была выполнена без разрешения
преступление преступного ущерба или умышленного ущерба», — сказал профессор Янг.
«Единственная разница заключается в том, как законы соблюдаются.
«Для того, чтобы действовать по своему усмотрению в отношении судебного преследования, должно быть какое-то эстетическое оправдание, а для многих людей в обществе маркировка не дает этого.»
Инфографика: Сотрудник города Ярра снимает бирки с витрины магазина в Коллингвуде, штат Виктория. (Новости ABC: Аллисса Отред.)Австралия нашла свое собственное граффити в «диком стиле»
Доктор Макдауэлл сказал, что некоторые из причин, по которым люди боролись с маркировкой, заключались в неразборчивости, размещении и рекламном повторении в нескольких местах.
«Это понятно с точки зрения передачи сообщения… размещение, то, как буквы работают вместе; использование материалов; поток… ни одна из этих вещей на самом деле не имеет смысла», — сказал он.
Но отрицать эстетическую ценность тегов — значит игнорировать 40-летнее развитие местных стилей рук. , региональные стили развивались, и в Мельбурне появился характерный «дикий стиль», дополненный пригородными вариантами
Характерные формы букв, нацарапанные вдоль железнодорожных путей, восходят к ранним авторам граффити.
«Вы можете поехать во Франкстон и увидеть стиль, который очень популярен в этом районе, а затем отправиться в Хертсбридж и увидеть совершенно другой стиль», — сказал член группы граффити MC, DB, TSF «Muscles».
«Это своего рода трайбализм, местничество.»
Инфографика: Художник-граффити в заброшенном поезде Hitachi на западе Мельбурна. (Новости ABC: Аллисса Отред)
(Новости ABC: Аллисса Отред)Получение права собственности на теги
Возможно, самым трудным аспектом тегов было их размещение.
С тех пор как самые первые метки появились в транспортной сети Нью-Йорка, маркировка бросила вызов законам о собственности.
«Если кто-то нанесет бирку на вашу стену, [он или она] увидит в ней что-то, что он имеет право изменить, и это право, основанное не на судебном приказе или документе о праве собственности, а на Правильно, исходя из близости, из того, что стена как бы примыкает к общественному пространству, улице, тротуару», — сказал профессор Янг.
Инфографика: ‘Jokes’, художник-граффити с запада Мельбурна.(Новости ABC: Аллисса Отред) Во многом как и реклама, граффити оказывает воздействие, и многие активные создатели тегов оправдывают то, что они делают, возмездием против распространения корпоративной рекламы.
«Это похоже на человеческую реакцию на бескомпромиссные корпоративные логотипы… которые нельзя пойти и купить в магазине», — сказал Кларк Аарон из Giant Productions, мельбурнского городского магазина товаров для искусства.
Профессор Янг согласился: «Одна из причин, по которой использование тегов является сложным или непопулярным и противоречивым, заключается в том, что оно ясно демонстрирует идею о том, что вам не нужно верить в такую корпоративную культуру, что вы можете создать свою собственную.»
Хотя мы обычно уделяем мало внимания рекламе, мы часто осуждаем теги как эгоистичные акты бессмысленного вандализма.
— выбирать элементы субкультур для продвижения своей продукции, маркировка — это то, что не продается. Фото: Забор с бирками в пригороде Мельбурна Балаклаве.(Новости ABC: Аллисса Отред)
Точно так же, как политические и протестные граффити, которые когда-то были распространены в наших городах, в сегодняшней культуре, основанной на изображениях, маркировка служит напоминанием о том, что в демократии всегда будет способ бросить вызов статус-кво, добиться известности или просто участвовать в анонимной публичной маркировке.
«В конце концов, это краска на стене; это не имеет значения. Вы можете закрасить ее и сделать ее снова скучной серой», — сказал художник граффити ZFG «Jokes».
Кредиты
- Репортаж: Тим Стоун
- Фото: Аллисса Утред
Thrill + Fury: The Art of the Tag — это новый документальный сериал из трех частей, созданный специально для ABC. доступны онлайн сейчас.
Эп. 1 Знай свой продукт. Эп 2 Вставать. Эпизод 3 Разрушение — это созидание.
Темы: уличное искусство, современное искусство, закон-преступление-и-правосудие, Мельбурн-3000
Первый опубликованный 04 февраля 2016 г. 11:11:08
.
